


Introduction
Epidemiology of chronic kidney disease
Chronic kidney disease (CKD) affects over 10% of the world’s population worldwide, amounting to over 800 million individuals [1]. With the advancement in the pharmaceutical industry and the increasing availability of dialysis, the healthcare expenditure used for the treatment of CKD has risen drastically in the past decades [2]. In 2022, between 4.90 and 7.08 million patients worldwide had end-stage kidney disease and need kidney replacement therapy [3]. Of note, CKD represents an especially large burden in low- and middle-income countries, which are least equipped to deal with its consequences [4].
CKD is more common among older individuals, women, racial minorities, and people with diabetes mellitus and hypertension. Over the past two decades, CKD has become one of the leading causes of death worldwide. This disease is one of the few non-communicable diseases that has seen an increase in associated deaths during this time period [1,5]. Effective means for the diagnosis, risk stratification, and monitoring of kidney disease are much needed.
Podocyte as the focus of kidney disease
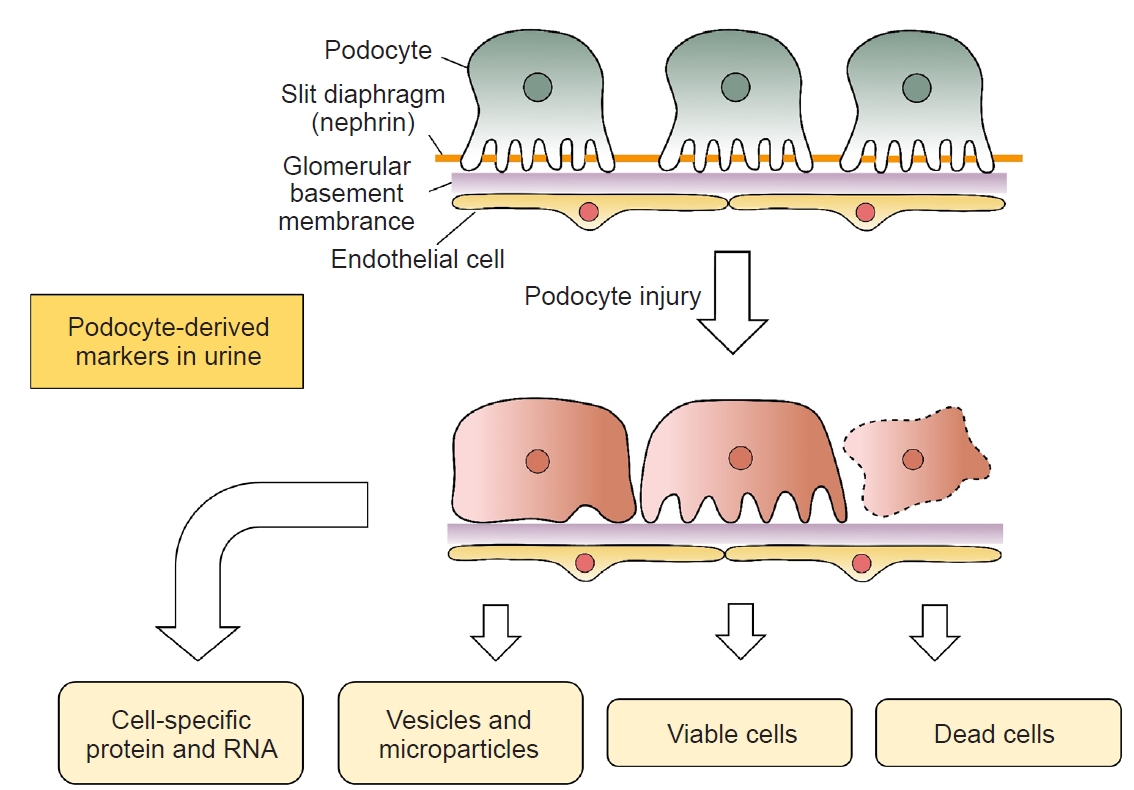
The primary function of the kidney is the excretion of metabolic wastes and excessive body water [6]. This objective is achieved by three processes: 1) filtration of plasma through the glomerulus; 2) reabsorption of useful substances via renal tubule; and 3) secretion of other metabolic wastes from peritubular capillary to the tubular fluid [6]. Although the kidney has a delicate three-dimensional (3D) structure with multiple cell types, the podocyte plays a central role in the maintenance of glomerular architecture and is the primary focus of many kidney diseases. Podocytes are terminally differentiated cells with a voluminous cell body that gives rise to primary processes, each of which has an extensive array of foot processes that affix to the glomerular capillary basement membrane (GCBM) and, together with the endothelial cells, from the glomerular filtration barrier [7]. The cell body of podocytes could be divided into three distinct compartments: the base, the top, and the slid diaphragm (SD) [8]. Specific cell membrane proteins are present in each compartment, and their interaction with the cytoplasmic cytoskeleton is responsible for the stability of podocyte structure and function [9].
Podocyte dysfunction plays a pivotal role in the pathogenesis of many kidney diseases. Germline mutations of podocyte-related genes result in kidney diseases, which typically present as steroid-resistant nephrotic syndrome and focal glomerulosclerosis (FGS). On the other hand, acquired podocyte dysfunctions, including a reduction in podocyte number or the density and fusion of podocyte foot processes, lead to the damage of glomerular filtration barrier and proteinuria [10]. Notably, many types of podocyte injury lead to a weakened interaction with GCBM, resulting in the detachment of podocyte and its cellular fragments to the urinary space, which may serve as biomarkers of kidney diseases (Fig. 1).
Podocyte damage and metabolic stress
A hot topic in podocyte research in recent years is the metabolic stress of podocyte during disease processes. Glycolysis and oxidative phosphorylation (OXPHOS) are the two major cellular pathways for energy production. Most cell types may switch between these two pathways in order to cope with any changes in energy demand, and podocyte is not an exception [11]. Podocytes are rich in mitochondria and heavily dependent on them for energy to maintain normal functions. A previous study found that mitochondrial OXPHOS contributes to over 50% of adenosine triphosphate production in mature podocytes [11]. Impairment of mitochondrial function can result in an energy crisis, oxidative stress, inflammation, and cell death. In diabetic kidney disease (DKD), mitochondrial pathways involved in podocyte injury include mitochondrial dynamics and mitophagy, mitochondrial biogenesis, mitochondrial OXPHOS and oxidative stress, and mitochondrial protein quality control. Mitochondria are dynamic organelles that respond to pathophysiologic cues by altering mitochondrial content, fusion, fission, mitophagy, and the unfolded protein response. Fission and fusion work together to maintain mitochondrial morphology, while mitophagy selectively removes damaged mitochondria from the network [12]. In DKD, excessive mitochondrial fission combined with decreased mitochondrial fusion is a typical feature of podocytes [13]. Additionally, the lack of proper mitochondrial turnover in the diabetic kidney is due to the inhibition of mitophagy [14,15]. Another key feature of mitochondrial dysfunction in podocytes is the reduced efficiency of mitochondrial biogenesis [15]. Under high glucose conditions, intracellular reactive oxygen species (ROS) production, mitochondrial DNA damage, and protein and lipid peroxidation are enhanced [16]. Furthermore, maintaining mitochondrial protein homeostasis is challenging due to the continuous exposure of mitochondrial proteins to mitochondrial ROS. Mitochondria cannot exist in isolation within a cell. They interact with the endoplasmic reticulum by forming mitochondrial-associated membranes (MAMs). The disruption of MAMs leads to abnormal intracellular Ca2+ influx, mitochondrial damage, and apoptosis [17]. The combination of the above factors results in podocyte injury and the progression of DKD.
However, mitochondria are mainly located around the podocyte nucleus and are scarce in the foot processes, probably because mitochondria are larger than the foot processes [18]. As a result, the energy expenditure of foot processes is largely supported by glycolysis, which probably explains the predominant expression of phosphofructokinase, the rate-limiting enzyme of glycolysis, in podocyte foot processes [11,18], and there were relatively low levels of tricarboxylic acid cycle intermediates in the glomerulus [18]. Recent evidence suggests that anaerobic glycolysis and fermentation of glucose to lactate are the primary sources of energy for podocytes. Additionally, anaerobic glycolysis maintains the glomerular filtration barrier independently of mitochondrial metabolism [18]. In diabetic mice, compromised glycolysis results in increased amino acid catabolism in podocytes [19]. In DKD, ornithine catabolism contributes to activating mammalian target of rapamycin (mTOR) signaling and cytoskeletal remodeling in podocytes. Therefore, inhibiting ornithine catabolism or mTOR signaling may help mitigate podocyte damage in diabetic mice [19]. Further studies showed that the blockade of glycolysis reduces the migration ability of podocytes [11], while cell-specific knockout of mitochondrial genes in podocytes does not lead to any major changes in the structure and function of podocytes [18].
Available evidence showed that the metabolic pathway of podocytes alters in response to cell damage. Under high glucose conditions, the cellular levels of glycolytic enzymes significantly increased, while that of tricarboxylic acid cycle and mitochondrial respiratory chain proteins significantly decreased [20], and similar changes were observed in kidney tissue samples of diabetes [21]. In streptozotocin-induced diabetic mice, podocyte-specific knockout of pyruvate kinase M2 (PKM2), another key enzyme of the glycolysis pathway, resulted in significantly aggravated kidney injury [21], indicating that increased glycolysis is part of the protective mechanism in response to podocyte injury.
It follows that the impairment of podocyte glycolysis promotes the development of DKD. Notably, activation of the renin-angiotensin system leads to PKM2 inhibition [22]. When podocytes are treated with angiotensin II, PKM2 expression rapidly decreases [22]. Specific deletion of PKM2 in mouse podocytes further exacerbates angiotensin II-induced podocyte damage, which was followed by foot process detachment and proteinuria [22].
Increased glycolysis leads to the formation of pyruvate and lactate, resulting in a certain degree of intracellular acidosis. Podocytes express the G protein-coupled receptor 81 (GPR81), the lactate receptor that is known to modulate the vascular endothelial growth factor A and N-methyl-D-aspartate receptor signaling cascades in podocytes, and regulate glomerular permeability and podocyte remodeling, respectively [23]. Lactate accumulation may therefore contribute to podocyte dysfunction in a diabetic milieu [24,25]. Since glomerular mesangial cells also express lactate transporters [26,27], it is possible that lactate accumulation in diabetic podocytes may also affect mesangial cell function.
Other metabolic disturbances in podocytes probably come secondary to the impaired glycolysis. Hyperglycemia leads to the accumulation of lipids in podocytes, which may accelerate the development of DKD, and this process may be aggravated in an environment with high angiotensin II level [28]. Adipose triglyceride lipase (ATGL), which is the first step in the hydrolysis of triglycerides, is important for maintaining the dynamic balance between lipid droplet storage and metabolism [29]. High-fat diet reduces ATGL expression and increases the accumulation of lipid droplets in podocytes, and the process can be reverted by promoting ATGL expression [30]. The depletion of ATGL in podocytes increases the intracellular level of ROS, promotes cytoskeleton redistribution, induces foot process fusion, increases the permeability of the glomerular filtration barrier, and promotes cell apoptosis [31]. The hyperglycemia-induced reduction in ATGL expression in podocytes is found to be the result of lactate accumulation, which activates the GPR81 receptor, with the downstream suppression of the cyclic adenosine monophosphate/protein kinase A pathway [31]. On the other hand, the facilitation of lipid beta-oxidation in human podocytes by the inhibition of acetyl-CoA carboxylase 2 alleviates glucose-induced insulin resistance via the sirtuin 1/peroxisome proliferator-activated receptor gamma coactivator-1 alpha pathway [32]. In addition, Luo et al. [19] showed that in a diabetic environment, impaired glycolysis of podocytes enhanced ornithine catabolism. Putrescine, the metabolite of ornithine catabolism, activated Rheb, thereby promoting the activation of mTOR signaling, which leads to cytoskeleton remodeling and fusion of podocyte foot processes [19].
Podocyte-specific targets as biomarkers
The major podocyte-derived molecules that are potential targets for biomarker development are summarized in Table 1.
Nuclear targets
Wilms’ tumor suppressor gene-1 (WT1) is a zinc finger-like transcription factor in the podocyte nucleus and is the most commonly used podocyte marker for histological study [33].
Slit-diaphragm protein complex
Components of the podocyte SD have been extensively studied as biomarkers of kidney diseases. The major targets in this group include nephrin, podocin, CD2-associated protein (CD2AP), and podoplanin [34]. Nephrin is a transmembrane protein with extracellular domains that form the core of the SD. It acts as a physical sieve barrier as well as a signaling scaffold [35]. Podocin connects nephrin to the actin cytoskeleton. CD2AP is a scaffolding molecule that interacts with nephrin and actin and is implicated in actin remodeling and membrane trafficking of podocytes [36]. Podoplanin is a small transmembrane glycoprotein in the SD compartment and has been implicated in the non-canonical signaling of mineralocorticoids [37].
Glomerular capillary basement membrane targets
The major protein components of the podocyte basal region are the α3β1 integrin and α-dystroglycan (α-DG). These proteins anchor podocytes to the GCBM and are critical for maintaining the orientation of podocyte as well as the integrity of the filtration barrier [38]. The α3β1 integrin is a transmembrane protein that connects GCBM with the cytoskeleton via α-actinin-4. The α-DG is a glycosylated protein that is connected to the cytoskeleton via utropin, and to the podocyte and GCBM via its interaction with laminin.
Targets at the top membrane
Podocalyxin is the major transmembrane protein on the apical side of podocyte. It is a negatively charged sialic acid protein and contributes to the charge barrier of the glomerulus [39]. It is connected to the actin cytoskeleton by ezrin and sodium-proton exchanger regulatory factor 2 (NHERF2).
Cytoskeleton targets
Several podocyte-specific proteins are linked to the actin cytoskeleton and maintain the 3D structure of the podocyte. The best-studied ones are synaptopodin and α-actinin-4. Both of them bind to the actin skeleton via interaction with the membrane-associated guanylate kinase with inverted orientation-1 (MAGI-1) [40]. Synaptopodin is only expressed in differentiating podocytes after they develop the foot processes, and it is often considered as the marker of mature podocytes [41].
Methods of study
Urinary podocytes
Intact podocytes are detectable in the urine of healthy people and patients with kidney diseases [44]. In fact, urinary loss is probably the major cause of podocyte depletion in CKD [45]. The traditional method for the identification of podocytes in urine involves cytospin and immunofluorescence study with specific antibodies (most commonly anti-podocalyxin antibody) [46]. The technique can be partly automated but has limited sensitivity and specificity because of cell debris contamination. With this technique, healthy people excrete less than 0.5 podocytes/mg creatinine, whereas patients with glomerular disease excrete up to 400 podocytes/mg creatinine [44].
In addition to the cytospin technique, podocytes could also be identified by the detection of podocyte-specific tryptic peptides with liquid chromatography coupled with tandem mass spectrometry (LC-MS/MS). Although the equipment cost is high, this technique is not operator-dependent and therefore highly reproducible [47].
The majority of urinary podocytes are viable when tested with propidium iodide exclusion and TUNEL staining [44]. In theory, a culture of viable podocytes ex vivo would improve the specificity of podocyte identification. However, podocytes do not normally proliferate in vivo, and special experimental conditions are required for the ex vivo culture [48]. This method has great value in translational study and therapeutic target identification but is too complicated for routine clinical use.
Urinary podocyte-derived fragments
Extracellular vesicles (EVs) are spherical membranous bodies released by various cell types [49]. In urine, EVs containing apical membrane and intracellular fluid are shed from all nephron segments, including podocytes, renal tubular epithelial cells, and uroepithelial cells. Urinary EVs may contain protein markers for kidney damage. In addition, EVs may contribute to intercellular communication within the nephron [50]. Previous studies showed that apical cell membrane fragments are shed from injured podocytes into the urine and could be identified as podocalyxin-positive granular structures by electron microscopy [51]. Urinary exosomes from podocytes could also be isolated by serial centrifugation, and microparticles quantitated by flow cytometry with annexin V antibody (which detects all microparticles), followed by antibody to podocalyxin or podoplanin (which specifically identifies podocyte-derived microparticles) [52,53]. Additional light-scattering analyses have been applied to determine the size of the microparticles [54]. With these sophisticated techniques, it has been shown that podocyte-derived microparticles are increased in mice upon exposure to high glucose [53], as well as in diabetic mice before the onset of albuminuria [54]. However, the application of these techniques to human kidney disease has not been explored.
In addition to measuring the number and size of podocyte-derived EVs and microparticles in the urine, there is a growing interest in measuring podocyte-specific molecules in urinary exosomes. For example, podocyte-derived signal transduction factors in urinary EVs have been found to be a promising candidate for the assessment of podocyte injuries [54]. However, their clinical applications remain under development.
Urinary podocyte-specific protein targets
The urinary levels of several podocyte-specific protein targets can be measured with ease, and their role as biomarkers of kidney disease has been studied extensively. In healthy adults, urinary podocalyxin and nephrin levels are generally detectable by enzyme-linked immunosorbent assay but not by traditional Western blotting [55]. Urinary nephrin level correlates with the severity of proteinuria [56] and kidney function [57]. Urinary podocalyxin level can also be measured by indirect immunofluorescence or flow cytometry [58], and it has been proposed as a marker for urinary podocyte count. Notably, urinary podocalyxin originated from the apical membrane of injured podocytes, and its level is increased in early kidney injury [51]. The major problem of using a urinary level of podocyte-specific protein targets as a biomarker is the confounding effect of urine concentration dilution. Urinary creatinine concentration is commonly used for adjustment, but this is only appropriate for free proteins in urinary supernatant but not for the levels in the cellular sediment.
Urinary podocyte-specific messenger RNAs
Measurement of urinary podocyte-specific messenger RNA (mRNA) level has been explored as the marker of intrarenal podocyte injury. Podocyte-specific mRNA level in urine is readily measured by real-time quantitative polymerase chain reaction (RT-QPCR). Two previous studies showed that urinary nephrin mRNA levels had a close correlation with urinary podocyte count [56,59]. In an animal study, urinary podocyte-specific mRNA levels also correlated with the rate of glomerular podocyte loss as determined by kidney biopsy [54].
Recent studies, however, have shifted the focus to derived urinary mRNA indices. In a rat model, the urinary podocin-to-nephrin mRNA ratio had a better correlation with the severity of histological progression than the absolute mRNA levels [60]. In healthy adults, urinary podocin mRNA-to-creatinine ratio (a marker of podocyte detachment), podocin-to-nephrin mRNA ratio (a marker of podocyte stress), and urinary podocin-to-aquaporin-2 mRNA ratio (a marker of relative podocyte injury versus tubular injury) correlated with the arterial blood pressure [61]. Glomerular injury is specifically associated with increased urinary podocin-to-aquaporin-2 and nephrin-to-aquaporin-2 mRNA ratios [62].
Urinary podocyte-specific micro-RNAs
Micro-RNAs (miRNAs) are short noncoding RNAs that regulate many biological pathways by targeting specific mRNAs [63]. Similar to mRNA, urinary miRNA level is easily measured by RT-QPCR [64]. Unlikely mRNA, miRNAs are resistant to degradation, which makes them promising biomarkers for clinical use [64]. A number of specific miRNA changes have been reported in kidney diseases [65]. For example, podocyte-specific loss of miR-23b, miR-24, and miR-26a was associated with rapidly progressive glomerular injury [66], and urinary miR-21, miR-124, and miR-192 levels correlated urinary nephrin, synaptopodin, and podocalyxin levels in diabetic patients [67]. In addition, there are studies that delineated the pathophysiological mechanism of miRNA-mediated podocyte dysfunction. Notably, miR-193a suppresses the expression of WT-1 and affects podocyte differentiation [68]. Similarly, miR-26a was the most abundantly expressed miRNA in the glomerulus of normal C57BL/6 mouse, and miR-26a was responsible for the regulation of podocyte differentiation and cytoskeletal integrity [69]. In human studies, urinary exosomal miR-26a levels were significantly higher in lupus nephritis than in healthy controls [69].
However, although many miRNA targets may be significantly altered in podocyte dysfunction, their urinary levels would unlikely to serve as podocyte markers. This is because podocyte contributes to only a small proportion of urinary miRNA. Any specific miRNA alterations observed in the podocytes may not be readily discernable in the urine, while any alteration in the urinary miRNA levels may reflect the pathological change in other renal cell types [70,71]. For example, miR-21 inhibits the expression of tissue inhibitor of metalloproteinase 3 (TIMP3) [72], miR-26a inhibits the expression of transforming growth factor beta 1 [73], miR-23b targets Ras GTPase-activating protein SH3 domain-binding protein 2 (G3BP2) [74], and miR-29c targets Sprouty homolog 1 (Spry1) [75]. Although these miRNA targets and their corresponding pathways are relevant for the pathogenesis of podocyte dysfunction, they may also be affected in other cell types in the kidney. Similarly, although urinary levels of miR-155, miR-663, and miR-1915 are significantly different between patients with FGS or minimal change nephropathy and healthy controls [76], it has not been shown that miR-155, miR-663, or miR-1915 specifically originates from podocytes.
Adjusting for urinary concentration
The key advantages and disadvantages of the above-mentioned methodologies are summarized in Table 2. An important methodological consideration for studying any urinary biomarker is how to adjust for the concentration-dilution effect of the urine. For the study of urinary electrolytes, the traditional methods are via the calculation of fractional excretion or the trans-tubular gradient [77]. Both of these methods require the concomitant measurement of plasma electrolyte level. For urinary biomarkers, a common method is to adjust for urinary creatinine concentration, which originates from the practice of meuring urinary protein- or albumin-creatinine ratio in chronic kidney disease [78]. However, this method is actually only suitable for molecules that are freely soluble in the urine, and therefore the concentration of which is truly affected by urinary concentration-dilution and could be adjusted appropriately by the urinary creatinine concentration. For molecular targets present in urinary sediment, the quantity in a centrifuged sample is not affected by any concentration-dilution effect of the urine, but by the duration of urine collection (i.e., if the rate of excretion is constant, an 8-hour urine sample should contain twice the amount of the molecule as compared to a 4-hour urine sample). A timed urine sample or expressing the result as a rate of excretion per hour would seem appropriate for this kind of molecular target—akin to the reporting of albuminuria [79]. For molecular targets in urinary microvesicles, the optimal method of adjustment is less well-defined, and it probably depends on the centrifugation protocol.
Urinary podocyte markers in diabetic kidney disease
There is a wealth of literature on the use of various urinary podocyte markers for the diagnosis and monitoring of DKD. Amongst all podocyte-specific protein targets, the most promising ones are podocalyxin and glycogen synthase kinase 3β (GSK3β). Podocalyxin level in urine was increased in diabetic patients before the onset of microalbuminuria, and therefore likely valuable for the early detection of DKD [80]. In patients with overt DKD, urine podocalyxin levels correlated with albumin-to-creatinine ratio [81,82]. On the other hand, GSK3β levels in urinary exfoliated cells were more accurate than albuminuria in discriminating diabetic patients with and without progressive renal impairment [83], this assay is cumbersome and may not be applicable to routine clinical practice. Our recent study showed that urinary phosphorylated GSK3β (p-GSK3β) level correlated with kidney function, but urinary total GSK3β, p-GSK3β level, its mRNA level, or the p-GSK3β/GSK3β ratio did not predict dialysis-free survival or the rate of kidney function decline [84].
As to other podocyte-specific mRNA targets, urinary mRNA levels of nephrin, podocin, synaptopodin, WT-1, and α-actin-4 were elevated in DKD [85], and they also correlated with urinary podocyte count, urinary nephrin level, albuminuria, and the severity of renal impairment [86]. More importantly, urinary podocyte-specific mRNA levels preceded the onset of microalbuminuria in diabetic patients [86], and the urinary podocin mRNA-to-creatinine ratio (a marker of podocyte detachment) predicted the subsequent rate of kidney function decline [87]. However, all of these studies have small sample sizes and the inclusion criteria are prone to selection bias. Further large-scale studies are necessary to validate their results before routine clinical application can be considered.
In addition to biomarkers specific for DKD, the urinary level of several podocyte-specific targets has been explored as biomarkers for CKD in general. For example, urine synaptopodin level correlated with kidney function in both diabetic and non-diabetic CKD, regardless of the degree of albuminuria [88]. However, not all kidney diseases have podocyte injury [89], and the association between podocyturia and proteinuria varied markedly in different diseases, indicating that urinary podocyte markers should be explored as disease-specific biomarkers [89].
Challenges and future studies
Although podocytes play a crucial role in the development and progression of DKD, the use of podocyte-associated molecules in urine as biomarkers of DKD is still in its early stages of development [90,91]. Before this technology can be applied in clinical practice, much work is needed to define the molecular target (or combination of targets) to be tested. Since the amount of mRNA and miRNA in urine is small, there is no reliable method to determine the gene expression library in urine [92,93], and identifying suitable targets is often a fishing exercise. Moreover, a urine sample contains separate compartments (i.e., conventional sediment, microvesicles, and truly cell-free supernatant), and podocyte-associated molecules (proteins, mRNA, and miRNA) are present in them at different concentrations [94,95]. Since the compartment to be tested determines the centrifugation protocol [50,96], it is a necessary but arduous task to define the suitable molecular target in each specific urinary compartment for clinical use.
In developing biomarkers for DKD, there is another theoretical aspect that is often overlooked. Specifically, two types of biomarkers can be identified: those that are specific to DKD, and others that are generic for any type of kidney damage. The type of biomarker identified depends on the study protocol and the selection of the control group [97]. It can be difficult to identify markers that are specific to podocyte injury or dysfunction, as opposed to markers of kidney inflammation or other pathological processes. However, if a reliable generic marker of kidney disease (a predictor of kidney function loss regardless of the underlying cause of kidney damage) were identified, it would have great clinical potential. Since podocyte loss is a central factor in the progression of many kidney diseases, we believe it would be beneficial to focus on the search for urinary biomarkers related to podocytes.
Taken together, although available studies provide preliminary ideas on the potential of podocyte-specific targets as biomarkers, there is much to do before they can be applied to clinical practice. While the published data focused on the potential of these biomarkers, they did not clearly define their clinical relevance or practical implications. It would be crucial to delineate the prospect of translation of these biomarkers into clinical practice, including the potential challenges and opportunities that may arise. Additionally, the validation of these biomarkers in large-scale studies is necessary to ensure their accuracy and reliability. Finally, it is important to consider the prospective impact of these biomarkers on patient management, including potential changes in treatment plans. By addressing these important considerations, we can gain a deeper understanding of the potential of podocyte-specific targets as biomarkers and their impact on clinical practice.
Conclusion
Podocyte injury plays an important role in the pathogenesis and progression of many kidney diseases. Urinary levels of podocyte-derived cellular fragments and podocyte-specific molecules may serve as biomarkers for the diagnosis and monitoring of kidney diseases. With the advance in our understanding of podocyte biology, the development of new technologies, and the increasing availability of existing ones, urinary podocyte markers are expected to have an expanding application. On the other hand, the identification of specific urinary podocyte markers may shed light on the pathophysiology of kidney diseases. Further research should focus on the standardization and automation of laboratory methods, as well as defining their added value to the available tests.
 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print






")









