


Introduction
Chronic kidney disease (CKD) is a serious public health problem worldwide [1]. Despite strict glycemic control and the use of antiproteinuric drugs (e.g., angiotensin-converting enzyme inhibitors [ACEIs] and angiotensin receptor blockers [ARBs]), the incidence of CKD continues to increase because of the increasing prevalence of diabetes mellitus, hypertension, and obesity, as well as population aging [2]. The prevalence of CKD has been reported as 14% in the United States [3], 10.8% in China [4], and 8.2% in South Korea [5]. A systematic review and meta-analysis of observational studies reported that the global mean prevalence of stage 3 to 5 CKD is 10.6% and that of stage 5 is 13.4% [6]. The recommended management of CKD includes nutritional therapy, treatment of albuminuria, cardiovascular risk reduction, avoidance of potential nephrotoxins, adjustments to the dosage of drugs such as antibiotics, and management of complications, including hyperkalemia, anemia, and metabolic acidosis [1].
As a common clinical cause of renal injury, obstructive nephropathy is usually caused by urinary tract stones, benign prostatic hyperplasia, and pelvic or ureteral tumors. It can occur at all ages from infancy to old age and eventually leads to renal fibrosis. In addition, it may lead to acute kidney injury (AKI) or CKD. Obstruction of the urinary tract decreases renal blood flow and the glomerular filtration rate (GFR), and hinders reabsorption of solutes and water, urine concentrate, and excretion of hydrogen and potassium. Renal fibrosis is the final pathway and histological outcome of most forms of progressive kidney disease, including long-term obstructive nephropathy. Unilateral ureteral obstruction (UUO) is an ideal reproducible experimental rodent model of progressive renal fibrosis [7]. Although the UUO model enables the study of the different stages of renal fibrosis development in an accelerated manner, the precise mechanisms underlying UUO-induced renal fibrosis are not fully understood. Oxidative stress plays a major role in tubular and/or cellular injury and initiates the overproduction of reactive oxygen species (ROS), interstitial inflammatory cell infiltration, activation of fibroblasts, upregulation of transforming growth factor beta 1 (TGF-β1), cell death (apoptosis, autophagy, ferroptosis, and necroptosis), deposition of the extracellular matrix (ECM), and renal fibrosis [8]. Oxidative stress also evokes mitochondrial damage and prolonged endoplasmic reticulum (ER) stress, both of which lead to apoptosis [9].
This review article summarizes the current understanding of the pathogenesis of UUO-associated renal fibrosis and discusses the current literature on the prevention and delay of the progression of this disease.
The unilateral ureteral obstruction model
In general, the UUO model is initiated in rodent animals, mainly rats and mice, by ligation of the left ureter, which leads to ureteral obstruction, hence the term “ureteral obstruction.” The reason for choosing the left ureter may be related to the lower position of the left kidney, which avoids damaging the liver. The ureter in the contralateral (right) kidney is not ligated and is called the nonobstructed or unobstructed kidney. Male animals are preferred because the presence of female reproductive organs complicates the surgical procedure. It is important that the animal’s body temperature is maintained and the kidney is kept moist during the surgery. In this procedure, the animal is anesthetized, and an incision is made in the flank or middle abdomen to expose the kidney and ureter. The left ureter is ligated with silk thread and the incision is sutured. The types of ligation are single ligation, double ligation, and two-place ligation and cut. Sham surgery is performed for the UUO procedure without ligation of the left ureter. The overall treatment protocol for UUO duration ranges from 3 days to 6 months, and most studies collect data on day 7 or 14. Regardless of the kidney under UUO, the contralateral kidney illustrates several alterations during chronic UUO, such as enlarged interstitial spaces and impaired mitochondrial function (after 14 days) [10], dilated distal tubules, cast formation, collagen deposition, and fibrosis (after 6 months) [11,12]. Interestingly, at day 7 and even day 14, the UUO does not influence renal function (e.g., serum creatinine, blood urea nitrogen, and cystatin C) [13,14]. This may be associated with the compensatory function of the contralateral kidney at the early stage of UUO. The various UUO animal models reported in the literature are listed in Table 1 [8,9,11,12,15–22].
Manifestations of unilateral ureteral obstruction-induced kidney pathology
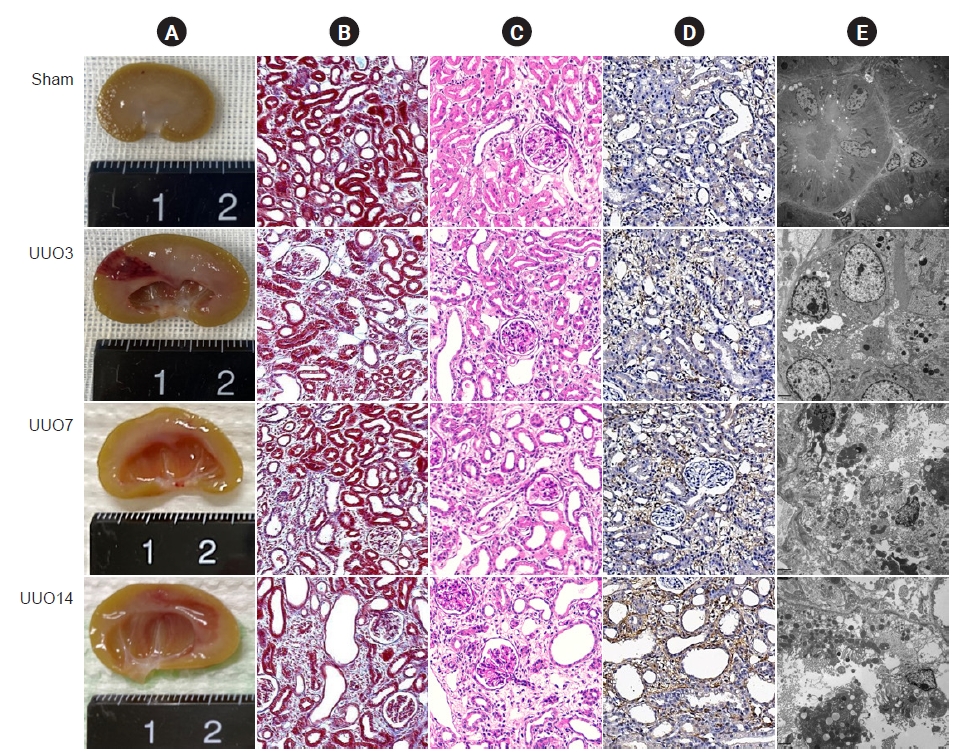
Images of the gross findings show that UUO results in enlargement of the kidney with hydronephrosis and pyelectasis, destruction of the renal pelvis, and an extremely thin kidney cortex (Fig. 1A) [7,23]. Pathology staining and transmission electron microscopy show that the major changes are confined to the tubulointerstitium and are characterized by necrosis, tubular vacuolization and atrophy, swelling of the tubular epithelium and interstitium, inflammatory cell influx, and fibrosis (Fig. 1B–D) [14,24].
Pathogenesis of unilateral ureteral obstruction-induced renal fibrosis
Renin-angiotensin system
Blockage of urine flow caused by UUO increases the hydrostatic pressure and impacts feedback to the distal and proximal tubules, which leads to mechanical stretching that damages the tubular cells and decreases the GFR. Both short-term UUO (as early as 6 hours) [25] and prolonged UUO (5 weeks) [26] activate the components of the renin-angiotensin system (RAS), such as expression of renin messenger RNA, ACE activity, and Ang II content, which contribute to renal fibrosis. These effects can be reversed by ARBs [27,28] or ACEIs [29]. Recently, our laboratory found that the angiotensin receptor neprilysin inhibitor LCZ696 alleviates renal fibrosis via ASK1-JNK-p38 MAPK-mediated apoptosis in the UUO model [30].
Oxidative stress
Oxidative stress plays a pivotal role in a wide array of kidney diseases through the overproduction of ROS [31]. In normal conditions, ROS production is strictly governed by the antioxidant system and ROS act as second messengers that help to regulate biological activity through oxidation of redox-sensitive proteins [32]. In response to pathological stimuli, the imbalance between the oxidant and antioxidant enzymes leads to an increase in the release of proteins involved in oxidation, which contributes to renal scarring [33]. Emerging evidence shows that oxidative stress contributes to renal fibrosis even in the early stage of UUO. Oxidative stress is manifested as lower activities of antioxidant enzymes such as catalase, superoxide dismutase (SOD), glutathione, and glutathione peroxidase in obstructed kidneys compared with sham-operated kidneys [34] and increased activity of oxidant enzymes, including nicotinamide adenine dinucleotide phosphate (NADPH) oxidases (NOXs), N(epsilon)-carboxymethyl-lysine, 8-hydroxydeoxyguanosine (8-OHDG), malondialdehyde, and 4-hydroxynonenal [24,35]. Pharmacological administration of antioxidant agents [14] or genetic knockout of cyclooxygenase 2 [36] or thioredoxin-interacting protein [37] alleviates renal fibrosis in UUO models.
Interstitial inflammation
Interstitial inflammation precedes ongoing fibrosis and is the predominant feature of renal fibrosis in UUO. The inflammatory response triggers a self-protective response against deleterious stimuli but sustained inflammation drives deposition of the ECM in the interstitium, which leads to irreversible fibrosis [38]. Numerous proinflammatory and profibrotic cytokines produced by renal tubular cells, myofibroblasts, and infiltrating macrophages are involved in the inflammatory response in UUO. Of these, macrophages are the major source of chemokines and cytokines such as monocyte chemoattractant protein 1, chemokine receptors, adhesion molecules, and TGF-β1 [39]. Chemokines and TGF-β1 also recruit macrophages, and this leads to a vicious cycle of inflammation [40]. Deficiency in NLRP3 or interleukin 36 (IL-36) attenuates renal fibrosis, as evidenced by blockade of ECM deposition, downregulation of NLRP3, pro-IL-18, pro-IL-1, and pro-caspase-1, and decreased inflammatory cell infiltration in UUO [41].
Fibroblast activation
Myofibroblasts belong to a heterogeneous superfamily derived from a variety of origins, including the epithelium through epithelial-to-mesenchymal transition, endothelia through endothelial-to-mesenchymal transition, local fibroblast or pericyte proliferation, and the macrophage-to-myofibroblast transition [42]. During renal fibrosis associated with UUO, activation of myofibroblasts is a key factor in the role of matrix-secreting cell types in ECM accumulation. About half of the myofibroblasts are created via the proliferation of local residential fibroblasts, 35% of nonproliferating myofibroblasts are created by differentiation of cells in the bone marrow, 10% come from the endothelial-to-mesenchymal transition program, and 5% come from the epithelial-to-mesenchymal transition [43]. TGF-β1, a member of the TGF-β superfamily, plays a critical role in this process via Smad-dependent and -independent signaling pathways [44].
Programmed cell death
Apoptosis (type I programmed cell death) is an active mechanism of cell clearance and plays an essential role in cell number regulation during development and organ homeostasis, and following insults [45]. In physiological conditions, apoptosis may be beneficial but can become deleterious if enough resident cells are lost [14]. The apoptotic program is classified according to the extrinsic and intrinsic pathways. Upon extrinsic stimulation, caspase-8 is activated by the ligation of death receptors that further activate downstream caspases by the extrinsic pathway. In the intrinsic (mitochondrial) pathway, proapoptotic Bax and Bak form pores in the mitochondrial outer membrane, which cause the release of apoptogenic factors (cytochrome C) and lead to stimulation of conformational modifications in the adapter protein Apaf-1 and subsequent activation of caspase-9 [46]. Conversely, Bcl-2 and Bcl-xL hinder pore formation via binding to Bax and Bak [47]. Overwhelming evidence has confirmed a pathological role of apoptosis in various kidney diseases as well as UUO [48].
Autophagy (type II programmed cell death) is an evolutionarily conserved, lysosome-mediated cellular process of degradation of damaged organelles, protein aggregates, and other macromolecules in the cytoplasm, and is involved in the regulation of cell death and proliferation [49]. The process of autophagy entails several consecutive procedures that produce autophagosomes and autophagolysosomes, and this process is controlled by a series of genes, such as autophagy-related Atg and LC3 [50]. Abnormalities in autophagy are implicated in a variety of kidney diseases [51]. In UUO, genetic ablation of Atg7 in distal tubules or pharmacological blockade of autophagy aggravates renal fibrosis, which suggests a protective role for autophagy [52]. However, this self-cannibalistic potential may be deleterious if excessive renal cells are lost. Livingston et al. [53] reported that persistent activation of autophagy worsens renal fibrosis in UUO mice with proximal tubule knockout of Atg7. Autophagy is also related to the upregulation of connective tissue growth factor and collagens, myofibroblast differentiation, apoptosis, mitochondrial dysfunction, and fibrosis [54]. Given its diverse actions, autophagy may serve a dual role as either a protector or contributor to kidney disease even within the same condition.
Intracellular organelles
The integrity of mitochondria and ER fitness play a pivotal role in organ pathophysiology [33]. In the kidney, mitochondria are more susceptible to oxidative stress because of arteriovenous oxygen shunting and their high oxygen consumption [55]. In kidney injury, oxidative stress causes mitochondrial ROS overproduction, which generates oxidative stress, alters the cell redox-sensitive signaling pathways, and results in inflammation and apoptotic cell death [31].
Sustained ER stress is cytotoxic to renal cells [33] and mitochondrial damage and ER stress are involved in the development of renal fibrosis in the UUO model [24,33]. In images generated using transmission electron microscopy, it is apparent that UUO destroys the mitochondrial architecture, as manifested by reductions in the number and size of mitochondria, vacuolization, mitochondrial deformation, and mitochondria divided into two daughter organelles (Fig. 2A). UUO is also closely associated with degranulation of ribosomes, disconnected and dilated cisternae, and peroxisome vacuolization in the rough ER, although the smooth ER remains almost normal in structure (Fig. 2B).
Necroinflammation
Necroptosis is a genetically regulated form of cell death controlled by receptor-interacting protein kinases 1 and 3 (RIP1 and RIP3) and downstream substrate pseudokinase mixed lineage kinase domain like (MLKL) [56]. Necrotized cells release danger-associated molecular patterns, which stimulate innate immunity to provoke the sterile inflammatory response via the ruptured plasma membrane. The sterile inflammation in turn aggravates necroptosis via tumor necrosis factor alpha (TNF-α) or interferon [57,58]. Therefore, the term “necroinflammation” refers to the reciprocal boosting of necrosis and sterile inflammation in an autoamplification loop [57,59]. Studies have suggested that RIP1-RIP3-MLKL-mediated necroinflammation is involved in the early stage of AKI [60]. However, Yi and OuYang [61] and Chen et al. [62] reported that necroinflammation participates in renal injury within 6 weeks in diabetic nephropathy (DN) and in the progression from AKI to CKD at 12 weeks. Of particular interest is that RIP3-MLKL signaling accelerates the inflammatory response by activating the NLRP3 inflammasome and thereby regulates the maturation of pro-IL-1β and pro-IL-18 [56]. We and others have reported that blockade of necroptosis with a specific RIP inhibitor (necrostatin-1 or GSK872) or RIP3 deficiency attenuates inflammation and fibrosis in UUO [13,24].
Renal fibrosis
Renal fibrosis is the final pathway of CKD progression that leads to end-stage renal disease requiring renal replacement therapy. Expansion and collagen fiber deposition within the tubulointerstitium are typical hallmarks of renal fibrosis. The major components of the ECM are collagens I, III, and IV, and fibronectin, and TGF-β1 may play an essential role. Consistent with the pleiotropic activities of this growth factor, emerging evidence suggests that TGF-β1 participates in the pathogenesis of a wide range of kidney diseases characterized by glomerulosclerosis and tubulointerstitial fibrosis [63]. In tissues, TGF-β is secreted as a biologically inactive complex requiring in vivo activation. The TGF-β1 inducible gene-h3 (βig-h3) encodes a matrix protein that is regulated by TGF-β. βig-h3 is regarded as an indicator of TGF-β1 bioactivity in renal injury and its expression is thought to indicate the extent of interstitial injury in the human [64] and rat kidney [65].
Congenital and epigenetic mechanisms
In addition to the above-mentioned factors, congenital and epigenetic mechanisms should also be considered. In clinical practice, congenital obstructive uropathy is a leading cause of pediatric renal failure associated with kidney dysplasia in infants and children. Most cases of congenital obstructive uropathy resulting from the posterior urethral valves are tightly linked to varying levels of injury to the bladder, ureter, and kidney. This clinical dilemma manifests as a reduction in nephrogenesis, glomerular maturation, and tubular cellular proliferation accompanied by a greater compensatory growth of the opposite kidney [66]. The mechanisms underlying congenital obstructive uropathy remain incompletely defined but may be associated with combinations of biomechanical signal transduction, growth factor expression, and responses of specific renal autoregulatory mechanisms. Therefore, numerous animal models of congenital obstructive nephropathy have been developed, such as the chick embryo, fetal rabbit, and opossum. Moreover, epigenetic mechanism refers to heritable alterations in target gene expression via post-translational modifications of protein complexes, namely histones, or DNA methylation without altering the underlying genetic DNA sequences. The post-translation protein modifications include acetylation, methylation, phosphorylation, sumoylation, ubiquitinylation, carbonylation and glycosylation, and DNA methylation. Of these, two major types of epigenetic regulations that have been implicated in the pathogenesis of kidney diseases are histone acetylation and DNA methylation. A growing body of evidence suggests that epigenetic modifications in gene expression participate in the process of renal fibrosis, inflammation, and epithelial-to-mesenchymal transition in CKD. Pharmacologic inhibition of histone deacetylase attenuates renal fibrogenesis by amelioration of inflammation and ECM gene transcription and restoration of bone morphogenetic protein 7 (BMP7) and Klotho protein in UUO [67,68]. By detecting their aberrant expression, epigenetic modifiers might serve as potential biomarkers for diagnosing UUO-related kidney disease, and they may also be used as therapeutic targets for the management of these diseases.
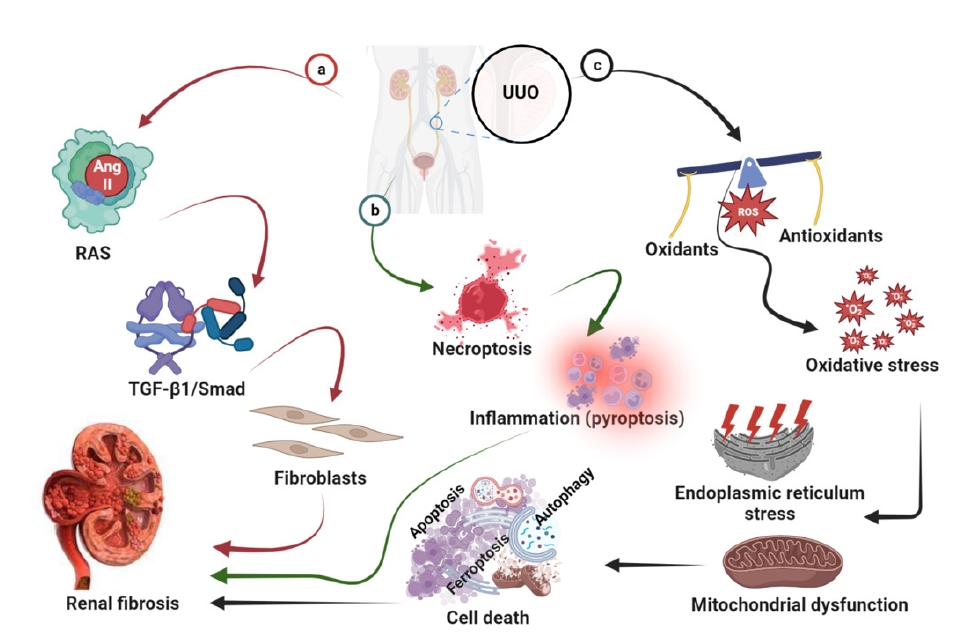
A diagram of the pathogenesis of UUO-induced renal fibrosis is shown in Fig. 3.
Management of unilateral ureteral obstruction-induced renal fibrosis
LCZ696
LCZ696 (valsartan/sacubitril) is a dual-acting angiotensin receptor neprilysin inhibitor. It is one of the new drug classes used for the management of hypertension and heart failure (HF) with reduced ejection fraction. The drug combines the angiotensin receptor blocker, valsartan, and the neprilysin inhibitor prodrug, sacubitril, in a 1:1 ratio in a sodium supermolecular complex [69]. Dual inhibition of neutral endopeptidase and Ang II by LCZ696 confers a better effect than each alone because it avoids reactivation of the RAS and angioedema [70]. The large PARADIGM-HF clinical trial reported that LCZ696 treatment caused a slower rate of decline in the estimated GFR, and improved cardiovascular outcomes compared with enalapril in patients with HF and CKD [71]. This concept is supported by animal studies of DN [72], subtotal nephrectomy [73], and UUO [30].
Vitamin E
Vitamin E is a fat-soluble vitamin also called alpha-tocopherol. Vitamin E has antioxidant potential and maintains intracellular integrity by protecting cells from oxidative stress [74]. In UUO, vitamin E ameliorates the progression of renal fibrosis by suppressing the TGF-β-Smad2/3 signaling pathway [75].
Curcumin
Curcumin is a natural phenolic compound extracted from the rhizome of the plant Curcuma longa (Zingiberaceae) that exhibits various biological activities. Previous studies have reported that curcumin improves renal dysfunction through regulation of the immune system, suppression of inflammatory mediators, oxygen free radicals, and apoptosis, and preservation of mitochondrial integrity [76]. A systematic review and meta-analysis of clinical trials found that curcumin supplementation has potential as a treatment for CKD [77] and diabetic kidney disease [78]. These beneficial effects of curcumin have also been reported in animal models of ischemia-reperfusion injury [79], DN [80], and UUO [81].
N-Acetylcysteine
N-acetylcysteine (NAC) is a modified amino acid that can reverse the toxic effects of acetaminophen overdose and prevent hepatic injury. The amino acid cysteine has a thiol side chain that can facilitate redox reactions and thereby exerts antioxidant activity [82]. Importantly, cysteine is needed for the synthesis of glutathione, which is a key intracellular antioxidant that protects against free radicals and other intracellular toxins [83]. Because of this activity, NAC may exert anti-inflammatory and antifibrotic actions in a variety of kidney diseases [84]. Shen et al. [85] reported that NAC attenuates the expression of collagen III and TNF, and renal fibrosis in UUO.
Sodium-glucose cotransporter 2 inhibitors
Sodium-glucose cotransporter 2 (SGLT2) inhibitors have been approved for the treatment of all stages of type 2 diabetes mellitus (T2DM). This class of drug directly blocks SGLT2 and thereby inhibits renal glucose reabsorption, boosts urinary glucose excretion, and effectively lowers hyperglycemia [86]. SGLT2 inhibitors afford better blood glucose control than other antidiabetic agents and have several advantages such as a low incidence of hypoglycemic episodes and lower body weight, and blood pressure [87]. Clinical trials have confirmed that SGLT2 inhibitors confer renoprotection and cardioprotection in patients with T2DM [88]. We have shown that dapagliflozin ameliorates renal fibrosis by inhibiting RIP1-RIP3-MLKL-mediated necroinflammation via Wnt3α/β-catenin/GSK-3β signaling in UUO rats [13].
Dipeptidyl peptidase 4 inhibitors
The first dipeptidyl peptidase 4 (DPP4) inhibitor was approved for the management of diabetes in 2006. This drug reduces blood glucose concentration primarily by impeding the degradation of glucagon-like peptide-1 and -2 (GLP-1 and GLP-2) and glucose-dependent insulinotropic peptide [89]. DPP4 is expressed predominantly in the glomeruli and the brush border of the S1 to S3 segments of the proximal tubule [90], whose actions include promotion of the absorption of cleaved dipeptides and regulation of the function of the sodium-hydrogen exchanger 3 in the proximal tubules. Inhibition of DPP4 may increase its bioavailability, thereby prolonging the half-life of insulin action. DPP4 inhibitors have been approved for treating T2DM as a monotherapy, add-on therapy, or combination therapy with other antidiabetic agents. Beyond glucose reduction, DPP4 inhibitors elicit renoprotective effects through either a GLP-1-independent proteolytic function or a nonproteolytic DPP4 function [91]. In a mouse model of UUO, the DPP4 inhibitor, LC15-0444, significantly decreased albuminuria, urinary excretion of 8-isoprostane, synthesis of several proinflammatory and profibrotic molecules, and renal fibrosis [92]. These findings suggest that DPP4 inhibitors have pleiotropic effects.
L-Carnitine
The quaternary ammonium compound L-carnitine (LC, L-β-hydroxy-γ-N-trimethylaminobutyric acid) acts as a key cofactor for the β-oxidation of fatty acids by facilitating the transport of long-chain fatty acids in the form of acylcarnitine esters across the mitochondrial membrane [93]. The derivatives of β-oxidation (two carbon molecules) are used by the Krebs cycle to produce adenosine triphosphate as a form of stored energy. LC limits free radical generation and thereby contributes to the maintenance of fatty acid β-oxidation in mitochondria and protects tissues from damage by the renewal of oxidized membrane lipids [94]. LC may possess antioxidant properties beyond its effects on lipid metabolism [95]. In our in vivo and in vitro studies, we have found that LC decreased UUO- or H2O2-induced oxidative stress (e.g., 8-OHdG, SOD, and NOX production) in the kidney and HK-2 cells, and anchored mitochondrial architecture, which subsequently inhibited apoptosis and fibrosis [14].
Coenzyme Q10
The coenzyme ubiquinone-10 (CoQ10) is an endogenous lipid-soluble compound and is regarded as an electron transporter in the electron transport chain (ETC) that transports electrons from ETC complexes I and II to complex III [96]. CoQ10 functions as a potent free radical scavenger that helps to secure the mitochondrial membrane potential and to protect protein from oxidation and DNA damage [97]. However, therapeutic applications of CoQ10 as a potential antioxidant are significantly impeded by its poor bioavailability because of its low solubility in aqueous media. For this reason, we developed water-soluble CoQ10 (CoQ10-W) to improve its bioavailability [98]. We found that CoQ10-W may provide more benefits than conventional CoQ10 by diminishing injury in the pancreas [99] and UUO-induced kidney [24]. Table 2 summarizes the management of UUO-induced renal fibrosis using clinical conventional drugs.
This review article elaborates on how to alleviate renal fibrosis using clinical conventional drugs in the adult UUO model. However, most cases of obstructive uropathy still require surgical intervention, such as diversion, stenting, and definitive surgery. Unlike adults, fetuses and infants require prenatal and postnatal interventions (e.g., shunt placement in utero).
Conclusions
Since its development in the 1970s, the UUO animal model has been greatly expanded and refined. This UUO model has several advantages over other models, including avoidance of exogenous toxins and uremia, shortened study duration, and low mortality rate in animals. Numerous investigators are exploring the mechanisms and prevention of renal fibrosis. This is of great clinical relevance because of the high prevalence of CKD worldwide. This review has focused on the current knowledge and therapeutic landscape relating to the mechanisms of UUO-induced renal fibrosis and the role of renoprotective agents such as conventional drugs in the management of renal fibrosis.

 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print






")









