



Introduction
Chronic kidney disease (CKD) is a common, global, progressive disease and a severe clinical problem associated with high mortality and morbidity [1]. Furthermore, there are no ideal drugs for the treatment or prevention of renal fibrosis, which is a major cause of the development and progression of CKD. In addition, renal fibrosis, regardless of its cause, is the final common stage in almost all CKDs [2]. Renal fibrosis is characterized by increased numbers of myofibroblasts, infiltration and accumulation of inflammatory cells, and excessive accumulation of extracellular matrix (ECM) components such as collagen and fibronectin [3].
In rodents, the unilateral ureteral obstruction (UUO) model is widely used to study progressive renal fibrosis because pathological features of CKD, such as tubular dilation, interstitial expansion, loss of tubular mass, leukocyte infiltration, tubular epithelial cell death, and the accumulation of myofibroblasts, are present in this model [4]. Pathogenically, hemodynamic changes are caused by mechanical stretching, apoptosis of epithelial tubular cells, oxidative stress, and inflammation, all of which contribute to progressive renal tubulointerstitial fibrosis [5]. The origins of renal myofibroblasts have been investigated over the last decade as the key reactive cells in renal fibrogenesis. Identifying the origins of renal myofibroblasts may elucidate the diverse characteristics and behaviors of myofibroblasts, which in turn could aid in the development of therapeutic strategies for the treatment and prevention of CKD [6]. Current evidence strongly suggests that collagen-producing myofibroblasts in the kidney originate from a variety of cellular sources, including bone marrow-derived cells, pericytes, resident fibroblasts, macrophages, and epithelial cells [7].
Diminazene aceturate (DIZE; C14H15N7.2C4H7NO3) is an aromatic diamidine that was developed more than six decades ago and which is currently marketed for the control of trypanosomiasis [8]. DIZE is also a well-known activator of angiotensin-converting enzyme 2 (ACE2), which converts angiotensin II to angiotensin (1–7) [9,10], and several reports have demonstrated that angiotesin (1–7) attenuates the development of cardiac fibrosis [11], renal fibrosis [12], pulmonary fibrosis [13], and hepatic fibrosis [14]. Besides being an ACE2 activator, the drug’s beneficial effects may be ACE2-independent in many pathological conditions [15]. Indeed, it has been suggested that DIZE exerts anti-inflammatory and antifibrotic activities in many human chronic disease models through angiotesin (1–7) independently of ACE2 expression/activity or angiotensin levels. For example, Rajapaksha et al. [16] demonstrated that DIZE protects against liver injury and biliary fibrosis by ameliorating proinflammatory cytokine levels, cellular oxidative pathways, and profibrotic cytokine expression without affecting ACE2 expression/activity or angiotensin levels. Similarly, cardiac fibrosis and diastolic dysfunction were ameliorated by DIZE treatment without affecting ACE2 activity [17]. However, the role of DIZE in kidney fibrosis and the underlying mechanism remain unclear. Therefore, we investigated the effects of DIZE on the progression of renal fibrosis after UUO in mice.
Methods
Surgical preparation of animals
All experiments were conducted using 8-week-old C57BL/6 mice (male and female) weighing 20 to 25 g each. The study was approved by the Institutional Animal Care and Use Committee of Pukyung National University (No. PKNUIACUC-2021-49) and was conducted in accordance with the Guide for the Care and Use of Laboratory Animals, published by the U.S. National Institutes of Health (NIH Publication No. 85–23, revised 2011). Mice were allowed free access to water and standard mouse feed. Animals were anesthetized with pentobarbital sodium (60 mg/kg body weight, intraperitoneal injection [i.p.]; Sigma-Aldrich) before surgery. To induce ureteral obstruction, the right kidney was exposed via a flank incision, the right ureter was completely obstructed using a 6-0 silk thread, and the incision was subsequently sutured [18]. Some mice were intraperitoneally injected with DIZE (15 mg/kg body weight, i.p.; Sigma-Aldrich) daily for 7 days after surgery. The dosage of DIZE was based on previous studies [10,19]. Body temperature was maintained at 36.5–37 °C throughout all surgical procedures using a temperature-controlled heating device (FHC, Inc.). Seven days after surgery, the kidneys were excised and snap-frozen in liquid nitrogen for molecular analysis or perfusion-fixed in 4% paraformaldehyde (Sigma-Aldrich) for immunostaining and histological studies.
Western blotting
Western blot analyses were performed as described previously [18] using the following antibodies: anti-α-smooth muscle actin (α-SMA, 1:10,000 dilution; Sigma-Aldrich), anti-phosphorylated SMAD3 (p-SMAD3, 1:2,000 dilution; Abcam), nuclear factor kappa B (NF-κB, 1:2,000 dilution; Cell Signaling), and Ly6G (1:2,000 dilution; eBioscience). Glyceraldehyde 3-phosphate dehydrogenase (GAPDH, 1:10,000 dilution; Bioworld Technology) was used as a protein loading control. ImageJ software 1.53e was used to quantify band densities.
Real-time polymerase chain reaction
Messenger RNA (mRNA) was extracted from kidney samples using TRIzol solution (Ambion). mRNA was reverse transcribed into complementary DNA using M-MLV reverse transcriptase (Promega). Faststart Universal SYBR Green Master Mix (Roche) was used as the quantitative real-time (qRT)-polymerase chain reaction (PCR) reaction mix. We used CFX Connect Real-Time PCR Detection System (Bio-Rad). Sequences of sense and antisense primers used in the murine qRT-PCR are as follows: 5’-TTGTACGGCAGTGGCTGAAC-3’ and 5’-AGTTGGTATCCAGGGCTCTCC-3’ for TGF-β; 5’-ACCTGCTGCTACTCATTCAC-3’ and 5’-TTGAGGTGGTTGTGGAAAAG-3’ for MCP1; 5’-CCAAGGGTTGACTTCAAGAAC-3’ and 5’-AGCGAGGCACATCAGGTACG-3’ for MIP2; 5’-CTGAAAGCTCTCCACCTC-3’ and 5’-TGCTGATGTACCAGTTGGGG-3’ for IL-1β; 5’-ACCTGGTAGAAGTGATGCCC-3’ and 5-GAGAAATCGATGACAGCGCC-3’ for IL-10; 5’-AGATGGATGTGCCAAACGTCCTCA-3’ and 5’-AATATGCGAAGCACCTTGGAAGCC-3’ for IL-4; 5’-TGGCTTTGCTCATCCGCTAT-3’ and 5’-CCAAGGTGATCCCAGGTGTC-3’ for CX3CL1; 5’-CTCCTCATCCTGCCTAAGTTCT-3’ and 5’-GTGCCGCTCTCGTTTACCTC-3’ for PAI-1; 5’-GATGGATGCTACCAAACTGG-3’ and 5’-GGAGAGCATTGGAAATTGGG-3’ for IL-6; 5’-AATTCCACTGAAGCTGGGCA-3’ and 5’-TCGGCATATGGGCTCCATTC-3’ for ACE2; 5’-ACCACAGTCCATGCCATCAC-3’ and 5’-CACCACCCTGTTGCTGTAGCC-3’ for GAPDH.
Sirius red staining and Masson’s trichrome staining
Paraffin-embedded kidney tissue sections were stained to detect collagen deposition using the Picro-Sirius Red Stain Kit (Abcam) and Masson’s trichrome stain [20]. For picrosirius red staining, deparaffinized sections were covered completely with picrosirius red solution for 1 hour. Samples were then rinsed twice with acetic acid solution (Sigma-Aldrich). For Masson’s trichrome staining, deparaffinized sections were refixed in Bouin’s solution for 1 hour at 56 °C, and sections were stained in Weigert’s iron hematoxylin working solution for 10 minutes. After washing, sections were stained in Biebrich scarlet-acid fuchsin solution for 10 to 15 minutes, followed by immersion in phosphomolybdic-phosphotungstic acid solution for 15 minutes. Picrosirius red or Masson’s trichrome-stained sections were then dehydrated in a graded dilution series of ethanol. Finally, sections were mounted on coverslips with Permount (Fisher Scientific). Micrographs were taken randomly of cortical areas in 200× microscope image fields using a Leica DM2500 microscope (Leica Microsystems GmbH). Areas of collagen accumulation in stained kidney tissues were analyzed using Fiji ImageJ software.
Immunohistochemical staining
Immunohistochemical (IHC) staining was performed to confirm the expression of α-SMA, a marker of fibrosis, infiltration of macrophages using an antibody against F4/80, a marker of macrophages, and CD206, an M2 macrophage marker. Oxidative stress was assessed by staining for 8-OHdG, a biomarker of DNA oxidative damage. Paraffin-embedded kidney tissue sections were rehydrated, followed by antigen retrieval, peroxide quenching, and blocking. Then, kidney sections were incubated with primary antibodies in a humid chamber overnight at 4 °C. Primary antibodies against F4/80 (1:100; Bio-Rad), α-SMA (1:400; Sigma-Aldrich), NF-κB (1:200; Cell Signaling), CD206 (1:200; Abcam), and 8-OHdG (1:500; Abcam) were used. Secondary antibodies were horseradish peroxidase (HRP)-conjugated goat anti-rat immunoglobulin G (IgG), HRP-conjugated goat anti-mouse IgG, or HRP-conjugated goat anti-rabbit IgG (Bethyl-Laboratories). Hematoxylin was used to stain nuclei. Sections were observed using a Leica DM2500 microscope. Micrographs of microscope image fields in cortical areas were taken randomly at 200× and 400× magnification. α-SMA-, F4/80-, CD206-, and NF-κB-positive cells were counted and recorded using a counting tool. The density of 8-OHdG was measured by Fiji ImageJ software as described in a previous study [21].
Terminal deoxynucleotidyl transferase dUTP nick-end labeling assays
Terminal deoxynucleotidyl transferase dUTP nick-end labeling (TUNEL) assays were performed using the Deadend Fluorometric TUNEL System Kit (Promega) according to the manufacturer’s instructions. Briefly, 4-μm kidney sections were deparaffinized and rehydrated. Sections were then incubated with TUNEL reagent mixture for 30 minutes at room temperature and then washed three times with phosphate-buffered saline (5 minutes for each wash). Sections were mounted on coverslips with an antifade mounting medium. Images of microscope image fields in cortical areas were obtained randomly at 200× using a Leica DM2500 microscope. TUNEL-positive cells were counted and recorded in five fields per kidney.
Histology (periodic acid-Schiff staining)
Kidney paraffin sections were stained with periodic acid-Schiff (PAS; Muto Pure Chemicals) stain according to a standard protocol. To determine morphological damage to tubular cells, five randomly chosen microscopic fields of the cortex region of each kidney were analyzed using the following scoring method: 0, no damage; 1, mild damage with rounding of epithelial cells and dilated tubular lumen; 2, moderate damage with flattened epithelial cells, dilated lumen, and congestion of lumen; and 3, severe damage with flat epithelial cells lacking nuclear staining and lumen congestion, as described previously [18].
Enzyme-linked immunosorbent assays to measure angiotensin (1–7) and angiotensin II
Blood was collected from the inferior vena cava using heparin sodium salt (20 IU/mL; Bioworld). Plasma was obtained by centrifuging the blood samples at 8,000×g for 5 minutes at 4 °C. Kidney tissue was dissolved in the radioimmunoprecipitation assay lysis buffer using homogenizer and kidney sample was obtained by centrifuging the kidney lysates at 16,000×g for 20 minutes at 4 °C and transferring supernatant to fresh tubes. Enzyme-linked immunosorbent assays (ELISAs) of plasma and kidney samples were performed using an angiotesin (1–7) ELISA Kit (Novus Biologicals) and angiotensin II ELISA kit (Cusabio) according to the manufacturers’ instructions.
Results
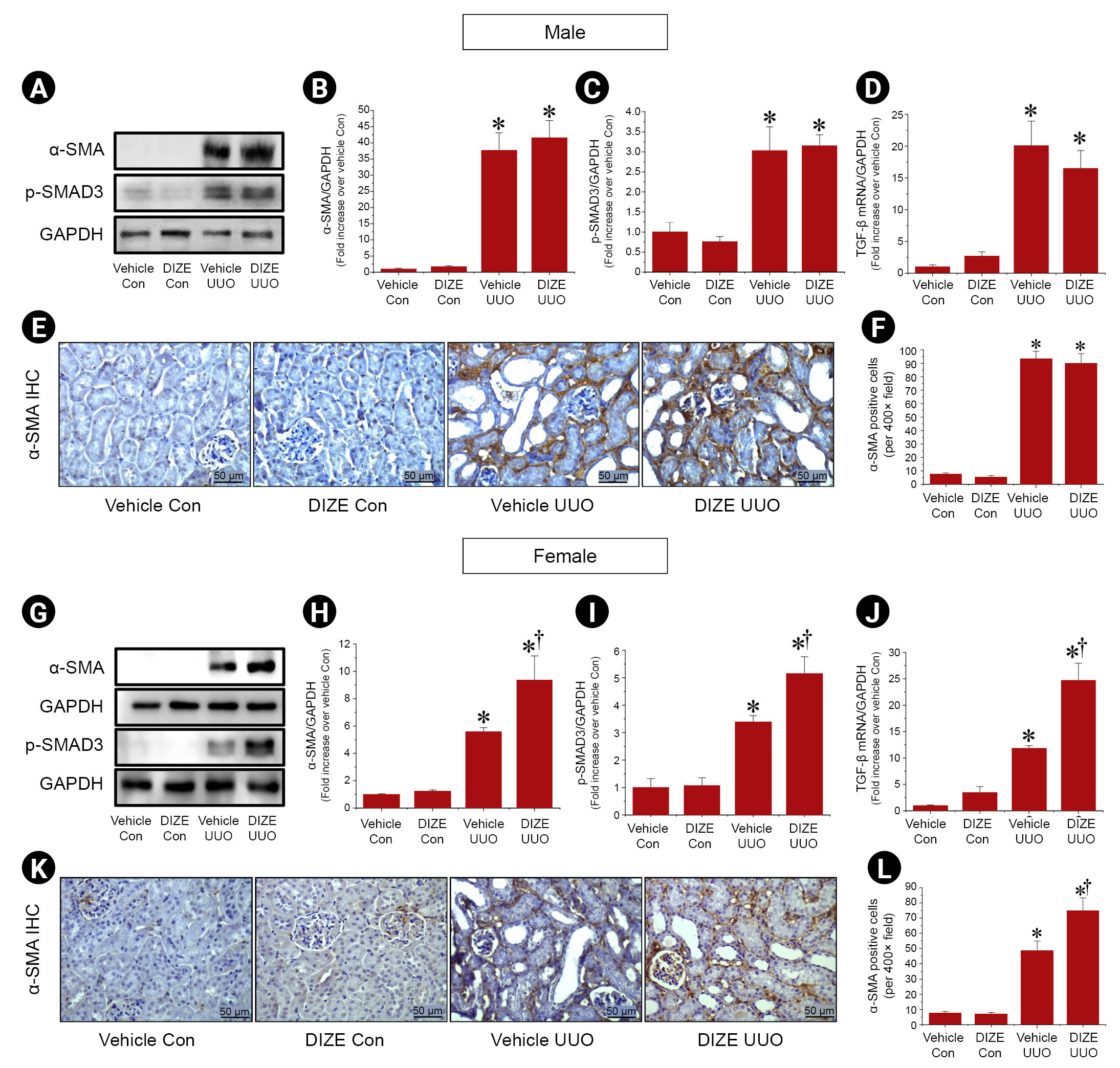
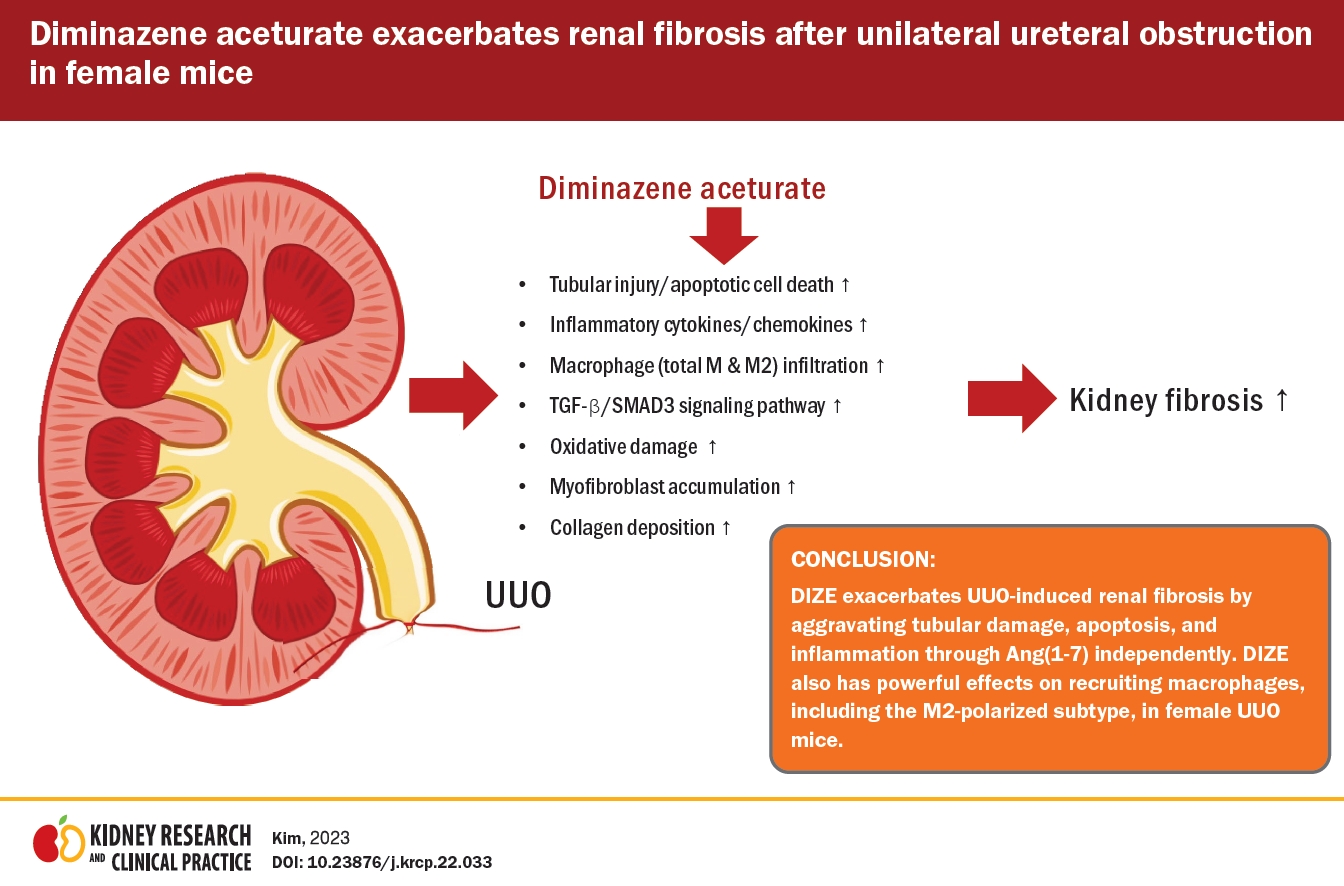
Diminazene aceturate exacerbates unilateral ureteral obstruction-induced renal interstitial fibrosis in female mice
To explore whether DIZE attenuates fibrosis in the UUO kidney, we first investigated α-SMA protein expression, a myofibroblast marker [22], by western blotting. UUO significantly increased α-SMA protein expression compared to that in control kidneys. DIZE treatment after UUO did not affect α-SMA protein expression or the number of α-SMA–positive cells compared to vehicle treatment in male mice (Fig. 1A, B, E, F). However, in female mice, DIZE further increased α-SMA protein expression and α-SMA–positive cells in UUO kidneys compared to the vehicle treatment (Fig. 1G, H, K, L). To further investigate the underlying mechanism, we examined whether DIZE affected the transforming growth factor (TGF)-β1/SMAD3 signaling pathway because TGF-β1 is a key mediator of fibrosis development and progression in CKD patients through SMAD3 phosphorylation [23]. SMAD3 phosphorylation and TGF-β1 mRNA expression were increased in female UUO kidneys compared to control kidneys, but DIZE treatment had no effect on SMAD3 phosphorylation or TGF-β1 mRNA expression in male mice (Fig. 1A, C, D). By contrast, SMAD3 phosphorylation and TGF-β1 mRNA expression were higher in DIZE-treated UUO female mice than in vehicle-treated UUO female mice (Fig. 1G, I, J). There were no significant differences in α-SMA expression, SMAD3 phosphorylation, or TGF-β mRNA expression between vehicle-treated and DIZE-treated control kidneys (Fig. 1). Contrary to our original hypothesis, these results indicate that DIZE exacerbates UUO-induced renal interstitial fibrosis in female mice but not in male mice, suggesting that sex may play a critical role in determining the effects of DIZE on renal fibrosis after UUO.
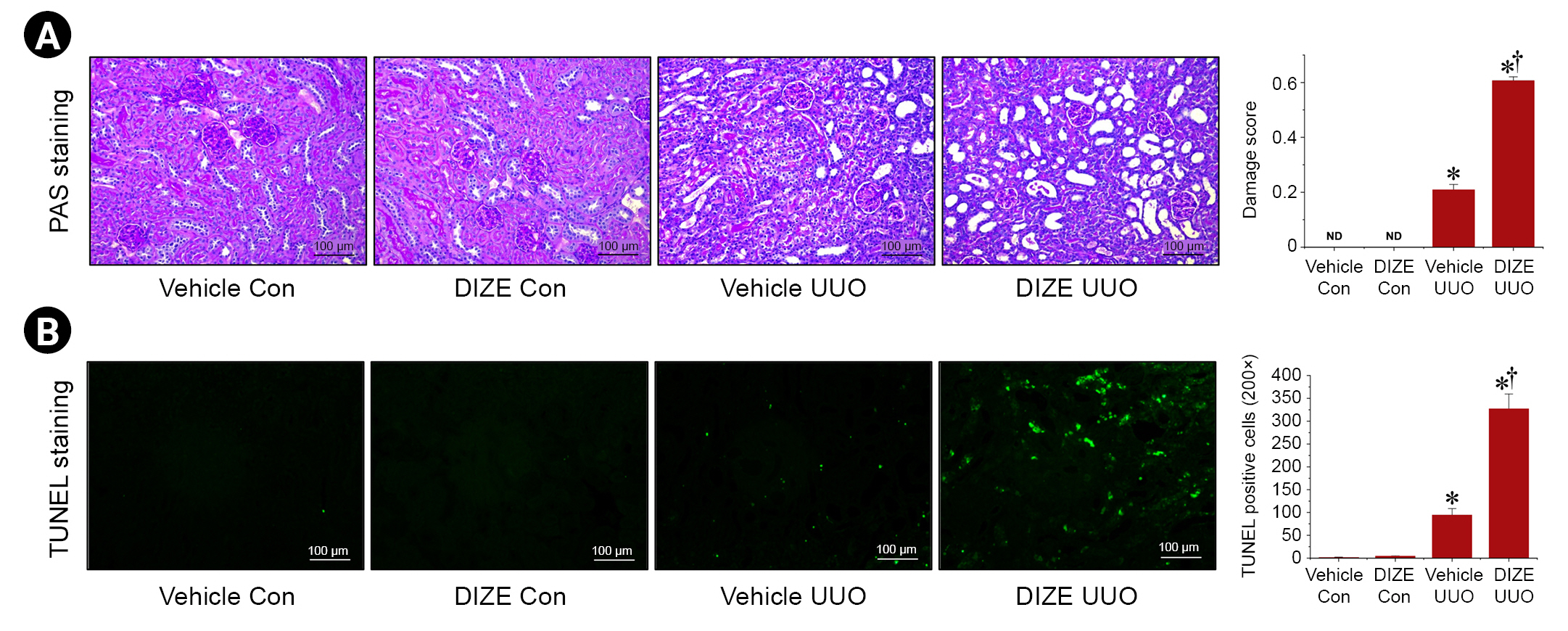
Diminazene aceturate aggravates unilateral ureteral obstruction-induced histological tubular damage and apoptotic cell death in kidneys
Next, we investigated if DIZE affects tubular cell damage and cell death. As expected, UUO-induced tubular damage is characterized by formation of proteinaceous casts and increased tubular dilation and congestion. DIZE treatment aggravated this damage in female mice (Fig. 3A). Moreover, we confirmed that UUO increased apoptotic cell death, as evaluated by TUNEL staining, and that DIZE treatment further increased apoptotic cell death in female mice (Fig. 3B). Taken together, these results indicate that DIZE exacerbates UUO-induced histological tubular damage and apoptotic cell death in the kidneys of female mice.
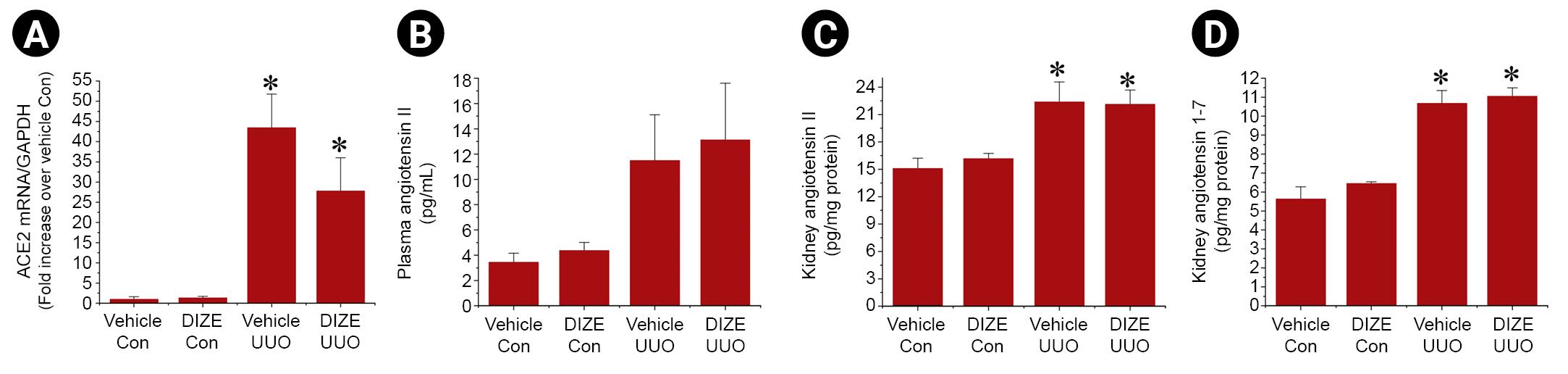
Diminazene aceturate treatment has no effect on angiotensin-converting enzyme 2 activity or angiotensin (1–7) and angiotensin II levels after unilateral ureteral obstruction
Because DIZE is a well-known activator of ACE2, which converts angiotensin II to angiotesin (1–7) [9,10], we assessed ACE2 activity by measuring angiotensin II and angiotesin (1–7) levels as well as ACE2 mRNA expression in kidneys (Fig. 4). Consistent with previous studies [24], we found that angiotensin II and angiotesin (1–7) protein levels, as well as ACE2 mRNA expression in kidneys, were significantly increased after UUO. However, DIZE treatment did not affect the UUO-induced changes in ACE2 mRNA expression or angiotensin II and angiotesin (1–7) protein levels in kidneys. These results suggest that DIZE exacerbates UUO-induced renal fibrosis independently of ACE2 after UUO in female mice.
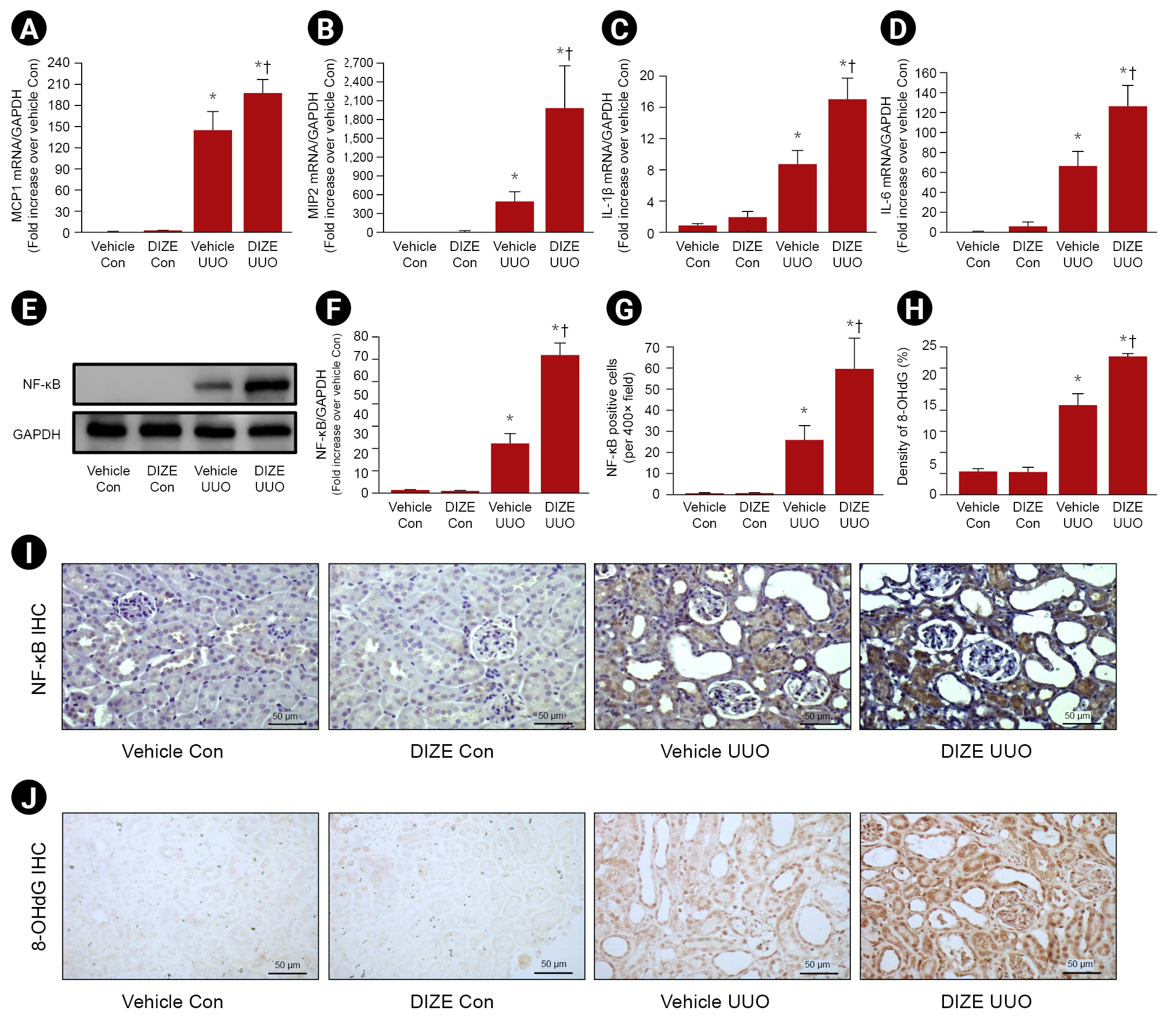
Diminazene aceturate aggravates inflammation and infiltration of macrophages in ureteral-obstructed kidneys
Given our finding that angiotesin (1–7) level had no relationship with DIZE-induced aggravation of renal fibrosis after UUO, we hypothesized that DIZE aggravates proinflammatory signaling and inflammatory cell accumulation in UUO kidneys, resulting in increased renal fibrosis. To test this hypothesis, we assessed the accumulation of inflammatory cells and levels of proinflammatory cytokines/chemokines. Proportion of F4/80-positive cells increased after UUO, and the increase in these cells was greater in DIZE-treated mice than in vehicle-treated female mice after UUO (Fig. 5A, B). We also confirmed that the protein expression of Ly6G, a neutrophil marker, was slightly but significantly increased after UUO. However, treatment with DIZE had no effect on Ly6G protein expression in UUO kidneys or control kidneys compared with that in vehicle-treated female mice (Fig. 5C). In addition, we also determined that macrophage infiltration and levels of cytokines/chemokines, monocyte chemoattractant protein-1 (MCP-1), macrophage inflammatory protein-2 (MIP-2), interleukin (IL) 1β, and IL-6 were increased after UUO, and that the increases in MCP-1, MIP-2, IL-1β, and IL-6 were greater in UUO kidneys of female mice treated with DIZE than in those of control mice (Fig. 6A–D). We also confirmed activation of the major proinflammatory NF-κB pathway using western blotting and IHC staining (Fig. 6E–G, I). UUO substantially increased the expression of NF-κB, and NF-κB expression was further increased by DIZE treatment. In addition, 8-OHdG, a biomarker of DNA oxidative damage, was increased after UUO, and DIZE treatment further increased levels of this biomarker (Fig. 6H, J).
Diminazene aceturate aggravates profibrotic M2 macrophage accumulation after unilateral ureteral obstruction
Acute tissue injury is caused by accumulation of proinflammatory M1 macrophages and the fibrotic response is driven by the persistent presence of profibrotic M2 macrophages [25]. M2 macrophages promote kidney fibrosis by producing excessive ECM and secreting profibrotic growth factors. As we had determined that DIZE aggravated the infiltration of total macrophages after UUO (Fig. 5), and because it was recently suggested that DIZE shifts macrophage polarization toward the profibrotic M2 phenotype in atherosclerotic lesions [26], we evaluated M2 macrophage accumulation by IHC staining. In addition, we measured levels of cytokines and chemokines involved in M2 macrophage survival and polarization. Using qRT-PCR, we found that mRNA expression of CX3CL1, IL-4, and IL-10, which are involved in M2 macrophage survival and polarization, increased after UUO, and DIZE treatment further increased the mRNA expression of these cytokines after UUO (Fig. 7A–C). In addition, the proportion of cells positive for CD206, an M2 macrophage marker [27], increased after UUO, and DIZE treatment further increased M2 macrophage accumulation after UUO (Fig. 7D).
Discussion
We originally hypothesized that DIZE treatment would protect against renal fibrosis after UUO. Our original hypothesis was based on the following previous findings: 1) DIZE is a well-known ACE2 activator that converts angiotensin II to angiotesin (1–7), which has antifibrotic and anti-inflammatory properties in diverse tissues, including the heart [11], kidney [12], lung [13], and liver [14]; 2) DIZE exerts anti-inflammatory and antifibrotic activities in many human chronic disease models via angiotesin (1–7) independently of ACE2 expression/activity or angiotensin levels; 3) the renin-angiotensin system (RAS), which is regulated by DIZE, is a critical regulator of renal fibrosis. To our surprise, we found that mice treated with DIZE had exacerbated renal fibrosis, inflammation, and M2 macrophage accumulation after UUO. Therefore, we suggest that DIZE has a detrimental role in renal fibrosis induced by UUO, rather than a protective role.
RAS regulates fluid balance, blood pressure, and maintains vascular tone. RAS is active in multiple tissues and organs, including the endothelium, myocardium, kidney, and brain. RAS consists of two pathways: the classical and non-classical pathways [9]. In the nonclassical axis, ACE2 converts angiotensin II into angiotesin (1–7), a heptapeptide that stimulates the Mas oncogene receptor (MasR) expressed in blood vessels. Numerous experimental studies have shown that angiotesin (1–7) exerts a vasoprotective role by increasing nitric oxide bioavailability, possibly through MasR, resulting in improved vasodilation [10]. In a diabetic mouse model, DIZE activated ACE2 and increased angiotesin (1–7) levels. These increased angiotesin (1–7) levels exerted a protective effect on diabetic mice by blocking the RAS system [10]. However, in the UUO mouse model, we and others [24,28] have found that angiotensin II and angiotesin (1–7) protein levels as well as ACE2 mRNA expression are significantly increased after UUO. DIZE treatment did not affect the UUO-induced changes in ACE2 mRNA expression or angiotensin II and angiotesin (1–7) protein levels in the kidney (Fig. 4). These results suggest that DIZE exacerbates UUO-induced renal fibrosis independently of ACE2 after UUO in female mice. Numerous studies have shown that kidney ACE2 activity as well as ACE2 mRNA and protein expressions are sex-dependent with higher ACE2 activity and expression in males than females [29–31]. In this study, we found that DIZE treatment after UUO did not affect renal fibrosis as assessed by α-SMA protein expression, SMAD3 phosphorylation, and TGF-β mRNA induction in male mice (Fig. 1A–F). Based on these findings, we speculate that the detrimental effect of DIZE on M2 macrophage-mediated renal fibrosis progression and the protective effects of DIZE on ACE2 activation may compensate for each other. However, how DIZE protects against renal fibrosis progression in UUO male mice remains to be defined in future studies.
Myofibroblasts are reactive cells with the combined characteristics of fibroblasts and smooth muscle cells. They are found in the presence of damage or under pathological conditions, such as cancer and human fibrotic diseases. α-SMA, which produces bundles of myofilaments termed stress fibers, is a myofibroblast-specific marker. Stress fibers connect the myofibroblast to the ECM; when the cell contracts, mechanical stresses are exerted on the matrix, triggering reorganization and wound closure throughout the healing process [18]. The majority of research has attributed this phenomenon to TGF-β, which performs its biological function by activating the SMAD3 signaling (p-SMAD3) pathway downstream and plays a vital role in the development of fibrosis. TGF-β activation controls a variety of cellular activities such as proliferation, apoptosis, differentiation, and inflammation. Furthermore, it stimulates fibroblast proliferation and ECM formation, resulting in profibrotic activity. In this study, we found that DIZE treatment markedly exacerbated UUO-induced renal interstitial fibrosis, activation of TGF-β/SMAD3 signaling, and myofibroblast accumulation (Fig. 1), indicating that DIZE plays a detrimental role in renal fibrosis by upregulating the profibrotic pathway.
Monocytes/macrophages are derived from bone marrow progenitors and enter the bloodstream. Circulating monocytes leave the bloodstream and enter tissues during homeostasis and inflammation. Following exposure to local growth factors, proinflammatory cytokines and microbial products induce the development of monocytes into macrophages [32]. In the early stages of CKD, macrophages infiltrate the renal cortex and increase the production of proinflammatory cytokines (IL-6, IL-1β, and TNF-α), reactive oxidative species such as superoxide anions and oxygen and nitrogen radicals, and matrix metalloproteinases; the increased production of these factors allows macrophages to migrate through basement membranes and interstitial ECM networks [33]. TNF-α attracts monocytes and macrophages to the tubulointerstitium of UUO kidneys by inducing upregulation of MCP-1 expression. In the present study, we found that macrophage production or levels of attracting cytokines/chemokines such as MCP-1, MIP-2, IL-1β, and IL-6 as well as macrophage infiltration were significantly increased by DIZE treatment compared to vehicle in UUO mice (Fig. 5, 6).
The monocyte-macrophage lineage can be activated by various stimuli and classified as either classically activated (M1) or alternatively activated (M2). Acute tissue injury is caused by proinflammatory M1 macrophages, and the fibrotic response is driven by the persistent presence of profibrotic M2 macrophages [25]. In CKD, accumulated M2 macrophages promote kidney fibrosis by producing excessive ECM and secreting profibrotic growth factors. For example, M2 macrophages secrete TGF-β, which contributes to the differentiation of progenitor cells into myofibroblasts and serve as a source of cytokines and growth factors with fibrogenic properties. They also secrete proteases, which participate in matrix remodeling [34]. Diverse factors can modulate M2 macrophage differentiation, recruitment, and survival. IL-4 and IL-10 polarize M2 macrophages by activating STAT6 and STAT3 via the IL-4 receptor (IL-4R) and the IL-10 receptor, respectively [32]. CX3CL1, also known as fractalkine, is expressed by renal tubular cells as a membrane-bound molecule, and CX3CL1 produced locally has the potential to promote CKD development by attracting CX3CR1-expressing M2 macrophages and fibrocytes into the kidneys [35]. A recent study demonstrated that DIZE increased α-SMA expression and promoted M2 macrophage polarization in an apolipoprotein E-knockout (apoE−/−) mouse model by stabilizing atherosclerotic lesions [26]. In this study, we also found that DIZE treatment aggravated M2 macrophage accumulation after UUO and increased the mRNA expression of CX3CL1, IL-4, and IL-10. In addition, the number of CD206 (M2 macrophage marker)-positive cells was increased by DIZE treatment after UUO (Fig. 7), suggesting that DIZE treatment exacerbates UUO-induced renal interstitial fibrosis, activation of TGF-β/SMAD3 signaling, and myofibroblast accumulation, at least in part, by M2 macrophage induction.
In summary, we demonstrated in this study that DIZE exacerbated UUO-induced renal fibrosis by aggravating tubular damage, apoptosis, and inflammation via angiotesin (1–7) independently of ACE2 expression/activity or angiotensin levels, rather than protecting against renal fibrosis after UUO in female mice. In particular, DIZE had a powerful macrophage-recruiting effect, including recruitment of the M2-polarized subtype, in UUO kidneys. Our findings suggest that therapeutic strategies with effects antagonistic to that of DIZE may help attenuate CKD and that treatment of kidney diseases should not focus solely on the previously reported therapeutic effects of DIZE.



 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print






")









