



Introduction
Primary cilia are nonmotile antenna-like cellular organelles that function as mechano- and chemo-sensors. The core of the primary cilium consists of microtubules with a 9 + 0 configuration anchored to the basal body. The length of the primary cilium changes dynamically in response to physiological and pathological stimuli. Defects in primary cilia cause a loss of flow-sensing ability and an osmotic stress response [1]. Recent findings have demonstrated that alterations in primary cilia length affect the functions of cells and organs and that aberrant primary cilia structure and function are associated with various diseases such as obesity, hypertension, diabetes, and polycystic kidney disease [2–5].
In the kidneys, primary cilia project into the tubular lumen and are directly exposed to urinary fluid flow, osmolality, and electrolytes such as sodium. Several studies have demonstrated that primary cilia length is associated with flow-sensing abilities [6,7]. We and others recently found that unilateral ureteral obstruction and unilateral nephrectomy change primary cilia length in renal tubule epithelial cells, both in the contralateral kidney and the remaining kidney [8–11]. Furthermore, we found that primary cilia length in the kidney varies during the injury and repair process [9–12]. Based on a clinical study, Verghese et al. [13] reported that cilia length was correlated with the function of the transplanted kidney. Similarly, evidence is accumulating that the length and composition of the primary cilia are associated with renal function [8–11]. However, how primary cilia respond to the intrarenal environment remains unclear.
Elongation and shortening of primary cilia depend on the assembly and disassembly of microtubules, which are composed of tubulin [14]. Acetylation of α-tubulin induces the assembly of microtubules in primary cilia, whereas deacetylation induces the disassembly of these microtubules. Studies have demonstrated that microtubule assembly and disassembly are associated with histone deacetylase 6 (HDAC6, a member of the class II histone deacetylases), which regulates tubulin acetylation [15–17]. Smith et al. [18] recently reported that inhibition of HDAC6 stimulates ciliogenesis in nonciliary pluripotent stem cell-derived endothelial cells, which then enables mechano-sensation by these cells. Furthermore, it has been reported that localization of aquaporin 2 (AQP2), a channel protein that plays an important role in concentrating urine, is regulated by microtubules [19,20], which are central organelles in the regulation of cilia length. Together, these data suggest that primary cilia are associated with kidney function. Supporting this, it has been reported that changes in HDAC6 expression and activity are associated with autosomal dominant polycystic kidney disease (ADPKD), a disease characterized by primary cilia defects [21].
Therefore, we hypothesized that primary cilia length is associated with urine concentration, including the conservation of water and electrolytes. To test this hypothesis, we investigated whether WD affected cilia length in kidney tubule epithelial cells and translocation of AQP2, and if so, whether blockage of tubulin assembly by HDAC6 inhibition affected cilia length and urine output. We found that water deprivation (WD) shortened cilia length in kidney tubule cells with concomitant concentration of urine, while HDAC6 inhibition blocked these WD-induced changes.
Methods
Animal
Ten-week-old C57BL/6 male mice (Koatech) were used in this study. All experiments were approved and performed in accordance with the approved guidelines of the Institutional Animal Care and Use Committee of Kyungpook National University, Republic of Korea (No. KNU-2022-0335). Mice either had free access to water (normal water intake, NWI) or were not allowed access to water (WD) for 24 or 48 hours. During WD, mice were given free access to standard mouse chow. Tubastatin A (a specific inhibitor of HDAC6 activation, 10 mg/kg body weight; Selleckchem) or 2% DMSO/saline (vehicle) was injected intraperitoneally each day starting from 2 days before WD until sacrifice. The dose of tubastatin was determined based on previous studies [22]. Kidney samples were obtained for biochemical and histological studies as described previously [23].
Scanning electron microscopy
Scanning electron microscopy (SEM) images were obtained using a scanning electron microscope (H-2500; Hitachi) as described previously [24]. Briefly, kidneys were perfusion-fixed with a fixing agent (0.5% glutaraldehyde and 0.5% paraformaldehyde) and then immersed in the fixing agent for 12 hours. Kidneys were post-fixed with 1% osmium tetroxide for 1 hour at 4 °C. After rinsing with 0.1-M phosphate buffer, kidneys were immersed serially in 25% and 50% DMSO for 30 minutes each. Kidneys were frozen rapidly on a metal plate by chilling with liquid nitrogen and then cracked using a scalpel and hammer. Cracked kidneys were thawed with 50% DMSO, washed thrice with 0.1-M phosphate buffer, and placed in 1% osmium tetroxide for 1 hour at 4 °C. Kidneys were transferred to 25% tannic acid for 2 hours at room temperature and placed in 1% osmium tetroxide for 1 hour at 4 °C. Kidneys were dehydrated using an ethanol series and isoamyl acetate. Dehydrated kidneys were subjected to critical point drying and then mounted.
Immunochemical and immunofluorescence staining
Kidney sections and fixed cells were immunostained as described previously [10]. For immunochemical staining, sections were stained with anti-AQP2 (Cat. No. AQP-002; Alomone Laboratories) antibody. Hematoxylin was used for counter-staining. For immunofluorescence, the following antibodies were used: anti–acetylated (ac)-α-tubulin (Cat. No. T7451; Sigma-Aldrich), anti–Na/K-ATPase (Cat. No. ab76020; Abcam), anti-AQP1 (Cat. No. AQP-001; Alomone Laboratories), and anti-AQP2 (Cat. No. AQP-002). To detect cell nuclei, DAPI was applied to samples. Images were captured using a Leica microscope (DM2500; Wetzlar).
Measurement of primary cilium length
Five to 10 images per kidney (five animals) were randomly captured under 400 magnification using a Leica microscope (DM2500), and then primary cilium length was measured in more than 100 cells using i-Solution software (IMT i-Solution Inc.). Cilium length was measured by tracing the cilium curvilinear line with several straight lines as instructed by the user’s guide for i-Solution. Primary cilium length in cultured cells (3–5 experiments) was measured in more than 50 cells. Cilium length was measured blindly by a person unaware of the grouping of the samples.
Bromodeoxyuridine incorporation assay
To assess cell proliferation, bromodeoxyuridine (BrdU, 50 mg/kg body weight; Sigma-Aldrich) was administered to mice every other day until sacrifice starting from 1 day before WD. BrdU was determined by immunohistochemical staining using anti-BrdU (Cat. No. MCA2060; Serotec) antibody.
Measurement of histone deacetylase 6 activity
HDAC6 activity was measured using an HDAC6 activity assay kit (Biovision Inc.) according to the manufacturer’s instructions.
Western blot analyses
Western blot analyses were performed as described previously [24]. Antibodies used were as follows: anti-proliferating cell nuclear antigen (anti-PCNA, Cat. No. m879; DAKO), anti–ac-α-tubulin (Cat. No. T7451; Sigma-Aldrich), anti–α-tubulin (Cat. No. T7451; Sigma-Aldrich), anti-AQP2 (Cat. No. ab3274; Merck Millipore), anti–E-cadherin (Cat. No. 610181; BD Bioscience), anti-GAPDH (Cat. No. NB300-221; NOVUS), and anti–β-actin (Cat. No. A2228; Sigma-Aldrich).
Membrane and cytoplasmic protein extraction
Membrane and cytoplasmic protein extraction in the kidneys was performed using the ExKine membrane and cytoplasmic protein extraction kit (Abbkine) according to the manufacturer’s instructions. Fractions were confirmed by western blot analysis using anti–E-cadherin (BD Bioscience) and anti-GAPDH (NOVUS) antibodies as markers of the cell membrane and cytosol, respectively.
Blood and urine biochemistry
Urine was collected using metabolic cages for 42 to 48 hours before mice were sacrificed. Blood was collected during sacrifice of mice. Urine and blood were subjected to biochemistry analyses. Hematocrit was measured using a hematology analyzer (scil Vet abc Plus, Allied Analytic, LLC.). Urine and blood osmolalities were measured using a cryoscopic osmometer (Osmomat 030-D; Gonotec). Urinary proteins were determined by Coomassie brilliant blue assay according to the manufacturer’s instructions.
Statistical analyses
All data were analyzed using GraphPad Prism 6 software (GraphPad Software Inc.). Results are expressed as means ± standard errors of the mean. The statistical significance of differences among groups was assessed using Student t test for comparison between two groups (Fig. 1–3) or two-way analysis of variance with repeated measures followed by Tukey multiple comparisons post hoc test for more than three groups (Table 1, Fig. 4–5). Differences were considered statistically significant at p < 0.05.
Results
Water deprivation decreases the length of primary cilia in kidney tubule cells
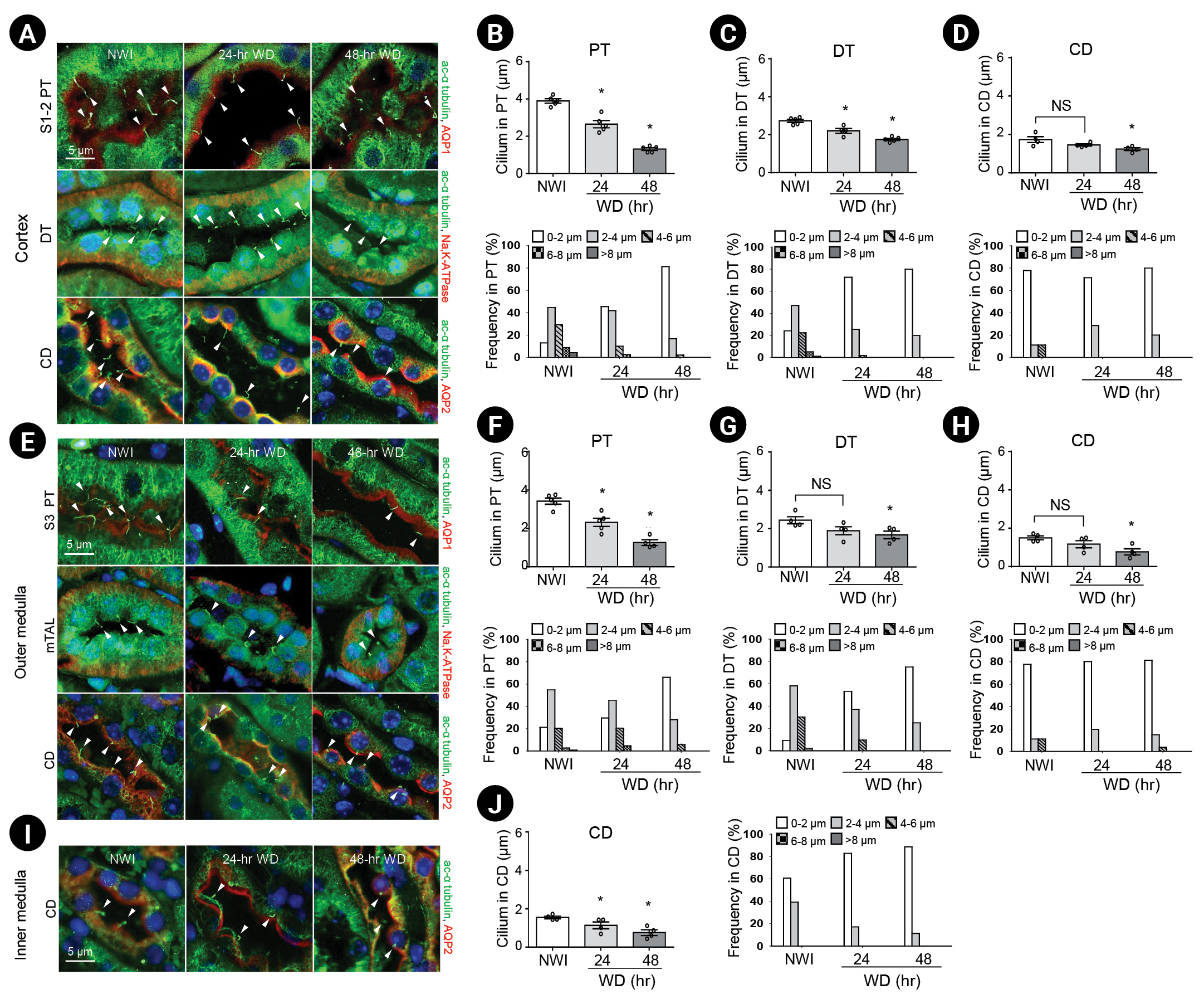
First, we determined whether WD affected the length of primary cilia in the kidney tubule epithelial cells of mice. When cells were observed using SEM, primary cilia were seen protruding into the tubular lumen of both WD and NWI mice (Fig. 1). Lengths of the primary cilia in WD mice were shorter than those in NWI mice (Fig. 1). When primary cilia were visualized by immunofluorescence using antibody to anti–ac-α-tubulin, primary cilia were observed in all renal tubule epithelial cells, with the exception of intercalated cells, which are known to not possess primary cilia [25]. Consistent with the SEM data, the lengths of primary cilia in kidney tubule cells in WD mice were shorter than those in the kidney tubule cells of NWI mice; the frequencies of short primary cilia increased gradually after WD. Primary cilia length of proximal tubular cells was the longest among tubules in NWI mice; primary cilia shortening after WD was greater in proximal tubules than distal tubules and collecting ducts (Fig. 2). These data indicate that WD induced the shortening of primary cilia.
To evaluate whether the decrease in primary cilia length was associated with cell proliferation (cells undergo deciliation before dividing) and/or cilia damage [11–13,26], we performed a BrdU-incorporation assay and examined PCNA expression to investigate proliferation, and also performed a urinary protein determination assay to look for cilia damage [10,12]. BrdU-positive cells were rarely observed, indicating that cell proliferation was not the cause of the decreased length of primary cilia (Fig. 3A, B). In addition, WD did not induce a change in PCNA expression in kidneys compared to NWI (Fig. 3C, D). Next, we evaluated whether WD-induced cilia shortening was due to the disruption of primary cilia and release of cellular contents into urine by determining urinary protein levels and ac-α-tubulin expression in the urine samples of both WD and NWI mice. The urine of WD mice contained higher amounts of proteins compared with the urine of NWI mice (Fig. 3E). Ac-α-tubulin in the urine of both WD and NWI mice was virtually undetectable by western blot analysis (Fig. 3F; the last lane in Fig. 3F represents urine collected from the kidneys with ischemia/reperfusion injury as a positive control). These data indicate that the WD-induced decrease in cilia length was not associated with either cell proliferation or cilia disruption.
Water deprivation increases histone deacetylase 6 activity and deacetylates α-tubulin in the kidney
Deacetylation of α-tubulin causes the disassembly of the microtubule axoneme, a central structure of primary cilia, leading to a decrease in the length of primary cilia, and α-tubulin is a substrate of HDAC6 [16,27]. Therefore, we investigated whether the WD-induced decrease in primary cilia length was associated with HDAC6. First, we investigated HDAC6 activity in the kidneys. HDAC6 activity was significantly higher in the kidneys of WD mice than NWI mice (Fig. 4A). Ac-α-tubulin expression was significantly lower in the kidneys of WD mice than in NWI mice, but level of total α-tubulin expression was similar in these two groups of mice (Fig. 4B–D), indicating that HDAC6 deacetylates α-tubulin.
Next, we investigated if tubastatin prevented WD-induced primary cilia shortening. Primarily, we investigated if tubastatin blocked HDAC6 activation. Tubastatin A was administered 2 days before WD on a daily basis until sacrifice. Tubastatin treatment significantly blocked the WD-induced increase in HDAC6 activity (Fig. 4A) and decrease in ac-α-tubulin levels (Fig. 4B–D). In NWI mice, tubastatin slightly reduced HDAC6 activity with an increase in ac-α-tubulin levels (Fig. 4B–D). Secondarily, we measured primary cilia length. Tubastatin almost completely blocked the decrease in primary cilia length induced by WD (Fig. 4E–K). However, tubastatin in NWI mice did not induce significant changes in primary cilia length (Fig. 4E–K). These results indicated that the WD-induced shortening of primary cilia was associated with HDAC6 activation.
Water deprivation-induced urine production is regulated by histone deacetylase 6
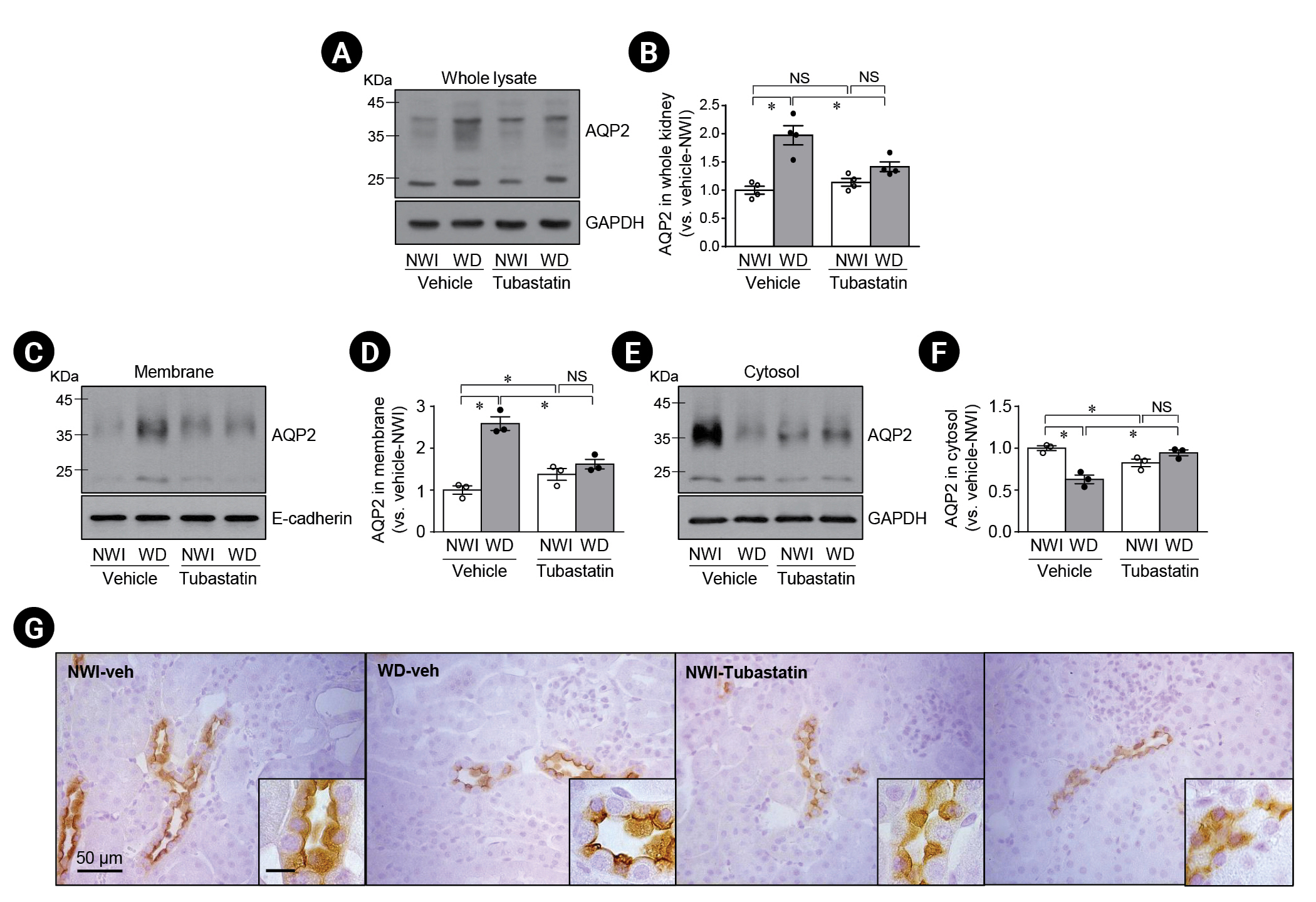
Since AQP2 is a critical player in the regulation of urine output and urine osmolality, we investigated if the localization and level of expression of AQP2 was associated with HDAC6 by quantitative and histological analysis, respectively. First, we determined AQP2 expression in whole kidney lysates. WD significantly increased AQP2 expression compared to NWI (Fig. 5A, B). Tubastatin treatment inhibited the WD-induced increase in AQP2 expression (Fig. 5A, B). In NWI mice, tubastatin slightly, but not significantly, increased AQP2 expression (Fig. 5A, B).
Second, we assessed AQP2 expression in membrane and cytosolic fractions from the kidney. WD increased AQP2 expression in the membrane fraction and reduced it in the cytosolic fraction (Fig. 5C–F). Tubastatin treatment significantly inhibited the WD-induced increase in AQP2 expression in the apical membrane fraction (Fig. 5C–F). In NWI mice, tubastatin slightly increased AQP2 expression (Fig. 5C–F).
Third, we determined the localization of AQP2 in the principal cells by immunohistochemical staining. Consistent with the western blot results, WD led to increased AQP2 expression at the apical plasma membrane with reduced cytosolic expression in principal cells (Fig. 5G). Tubastatin blocked the WD-induced membrane localization and increase in AQP2 protein expression (Fig. 5G).
Lastly, we investigated whether primary cilia length was associated with urine output and osmolality. Urine output from WD mice was significantly less than that from NWI mice and urine osmolality was higher in WD mice than in NWI mice (Table 1). However, there were no significant differences in plasma osmolality or hematocrit between WD and NWI mice (Table 1). Tubastatin injection in WD mice significantly increased urine output and decreased urine osmolality (Table 1). Tubastatin injection in NWI mice induced slight, but not statistically significant changes in urine output and osmolality compared with vehicle injection (Table 1). Plasma osmolality was not significantly affected by WD or tubastatin administration (Table 1). Tubastatin did not induce significant changes in the amount of water intake in NWI mice (Table 1). These data indicate that WD facilitates the apical localization of AQP2 and an increase in AQP2 expression through HDAC6 activation, consequently leading to concentrated urine production. Furthermore, tubastatin inhibits WD-induced apical localization of AQP2.
Discussion
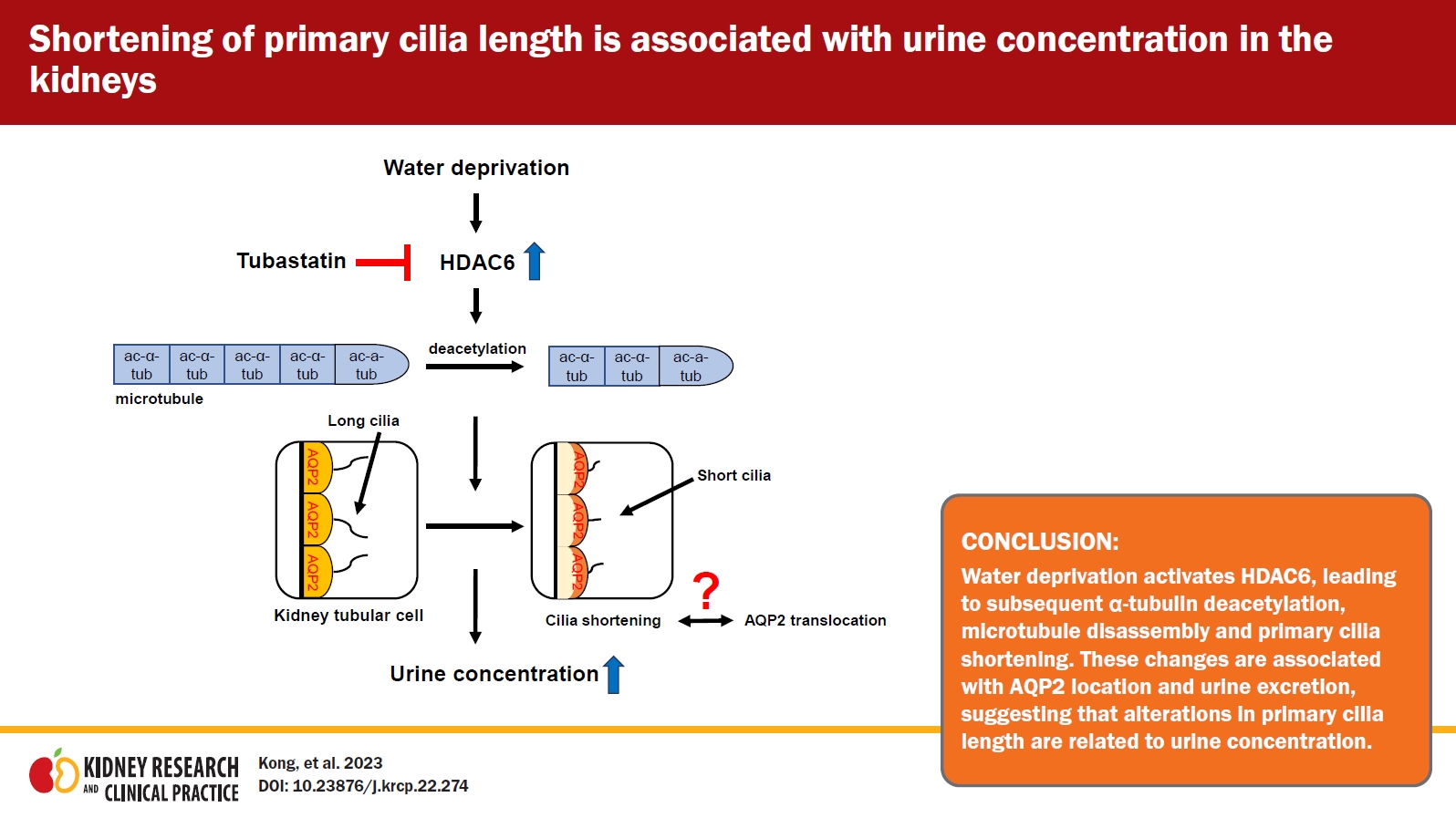
To the best of our knowledge, this is the first study to demonstrate that WD shortens primary cilia length in kidney tubule cells and that this is mediated by HDAC6 activity; HDAC6 inhibition impaired the water intake-associated changes in urinary output and osmolality as well as AQP2 localization. These data suggest that shortening of primary cilia length after WD is associated with urine concentration, and that, since HDAC6 inhibition impairs those kidney responses to water intake changes, HDAC6 is involved in this process. Our data provide insights into how primary cilia length is altered dynamically in response to physiological, stimuli, how this alteration in length is associated with kidney function, and how kidney tubule cells regulate primary cilia length.
Kidneys maintain water and osmolality balance through the excretion of urine. When water intake decreases, kidneys excrete concentrated urine via regulation of transporters and/or channels in kidney tubular epithelial cells. When water intake changes, renal tubule cell primary cilia are directly exposed to changes in fluid flow and osmolality and may respond to those changes. As expected, we found that in WD mice compared to NWI mice, there was a 1.82-fold increase in urine osmolality with decreased urine output. Unlike urine osmolality and urine output, hematocrit and plasma osmolality after WD tended to increase but not by statistically significant amounts. This indicates that plasma osmolality and hematocrit are tightly maintained within certain ranges under water-deprived conditions, with relatively minimal changes compared with urine volume. For these reasons, 48 hours of WD did not cause a significant imbalance in body water balance, and the body water balancing system compensated for this level of WD. Similarly, Brazzuna et al. [28] reported that 6 days of dehydration did not change hematocrit levels due to restriction of plasma water loss. de Fost et al. [29] reported that WD caused the release of arginine vasopressin into the circulation to preserve water and restore normal plasma osmolality. However, contrary to our results, Bekkevold et al. [30] reported that 48 hours of WD increased plasma osmolality by about 4% in mice.
In the present study, we observed that WD caused a decrease in the length of primary cilia, that the cilia length shortening induced by WD was associated with increased HDAC6 activity, and that this shortening was blocked by HDAC6 inhibition. Furthermore, tubastatin blocked WD-induced shortening of cilia, increased urine output, and decreased urine osmolality. In addition, we found that the length of primary cilia in proximal tubular cells was the longest among the tubules of NWI mice and that WD-induced primary cilia shortening in proximal tubules was greater than in distal tubules or collecting ducts. We speculate that the greater WD-induced shortening of cilia in proximal tubular cells may be due to the greater reduction in ultrafiltrate volume and flow compared to collecting ducts (or distal tubules) and the longer basal cilia length before WD in proximal tubular cells. Analysis of HDAC6 activity in each tubular segment may provide more insight into the link between HDAC6 activity and cilia length.
In previous studies, we found that the cessation of urine flow induced by ureteral obstruction resulted in a decrease in cilia length in renal tubule cells [8], whereas an increase in urine flow by unilateral nephrectomy led to elongated primary cilia in the remaining kidney [9]. In addition, we found that kidney tubule cells contained primary cilia of various lengths following a kidney insult such as ischemia/reperfusion injury, which induces dramatic changes in glomerular filtration rate and fractional excretion of electrolytes [23,31]. In addition, studies have reported that primary cilia are functionally involved in flow sensing [32,33]. Gilmer et al. [34] reported that the flow rate in proximal tubules is the greatest among renal tubules. Furthermore, it has been reported that osmolality-sensing proteins are localized on the membrane of cilia and that cilia are required for the response to osmotic stress [35]. Therefore, our data indicate that fluid flow and/or urine osmolality may cause alterations in cilia length and, conversely, that alterations in cilia length may contribute to urine output and osmolality. Therefore, we speculate that the cilia length changes and HDAC6 activation observed in WD-mouse kidneys may be normal responses of kidney cells and critical for the adaptation of kidneys to kidney burdens such as excessive hydration, leading to maintenance of body water balance.
We recently found that a decrease in the length of primary cilia was associated with deciliation and resorption of cilia [10–12,36]. Therefore, to define the involvement of cell proliferation and deciliation in dynamic changes in cilia length, we performed a BrdU incorporation assay and PCNA expression assay in kidney tissue and investigated the presence of ciliary proteins in urine. In previous studies, we reported that disrupted cilia in the kidney tubular epithelial cells were excreted into the urine [10–12,36]. In the current study, we found that WD-mouse urine contained greater amounts of protein than NWI mouse urine, but ac-α-tubulin was undetectable in the urine samples of both WD and NWI mice by western blot analysis. This indicates that primary cilia were not disrupted by WD. Furthermore, WD did not induce significant changes in the number of BrdU-positive cells or PCNA expression. Taken together, these data indicate that changes in WD-induced primary cilia length are not caused by either cell proliferation or deciliation.
The assembly and disassembly of microtubules by α-tubulin acetylation and deacetylation are associated with an increase and decrease in cilia length, respectively [10–12,26]. Recent studies have shown that HDAC6 serves as a regulator of primary cilia length; HDAC6 activation and overexpression induce deacetylation of α-tubulin and a subsequent decrease in primary cilia length [18,27,37]. Similarly, in the present study, we found that WD increased HDAC6 activity and led to α-tubulin deacetylation without a significant change in the total amount of α-tubulin expression. These data suggest that the shortening of primary cilia length induced by WD is associated with disassembly of microtubules and that this change may be an adaptive response to WD. Supporting this, inhibition of HDAC6 activation via tubastatin in water-restricted mice restored the ratio of ac-α-tubulin to total α-tubulin to that seen in NWI mice.
Microtubules are known to play an important role in the cellular localization and expression level of AQP2, a water channel that is essential for urine concentration [19,20]. Therefore, to define the functional association between cilia length changes and HDAC6, we evaluated WD-induced AQP2 localization, because the apical localization of AQP2 is critical for the generation of concentrated urine formation [38]. As expected, WD resulted in the movement of cytosolic AQP2 to the apical plasma membrane, and tubastatin blocked the apical localization of AQP2 following WD. Furthermore, WD increased AQP2 expression in the kidneys. Furthermore, tubastatin increased the WD-induced changes in urine output, osmolality, and cilia length to levels similar to those observed in control mice. These data indicate that WD regulates urine concentration through the apical membrane localization and de novo production of AQP2 and that this is at least partly regulated by HDAC6 activation. Smith et al. [18] recently reported that inhibition of HDAC6 activates cilia formation in human-induced pluripotent stem cell-derived endothelial cells lacking cilia and restores their mechano-sensing ability. Rymut et al. [39] reported that HDAC6 inhibition reversed the cystic fibrotic phenotype, with lowering of ac-α-tubulin levels. Ke et al. [21] reported that HDAC6 expression and activity were low in kidney tubule cells in ADPKD, a disease associated with defects in the primary cilia of kidney tubule cells. Furthermore, in a previous study, we found that inhibition of microtubule dynamics by taxol treatment during recovery delayed the restoration of kidney functional damage [40]. Therefore, we speculate that alteration of primary cilia length regulated by HDAC6 is associated with urine concentration.
In summary, although several limitations, including determining if there is a causal relationship between primary cilia and urine concentration remain, our data show for the first time that changes in primary cilia length are, at least in part, associated with urine concentration, suggesting that primary cilia length regulated by HDAC6 is involved in urine concentration and the maintenance of body water balance.



 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print






")









